This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

17 Q&As




Facts about Dual specificity tyrosine-phosphorylation-regulated kinase 1A.

Modulates alternative splicing by phosphorylating the splice factor SRSF6 (By similarity). Has pro-survival function and negatively regulates the apoptotic process.
| Human | |
|---|---|
| Gene Name: | DYRK1A |
| Uniprot: | Q13627 |
| Entrez: | 1859 |

| Belongs to: |
|---|
| protein kinase superfamily |

dual-specificity tyrosine-(Y)-phosphorylation regulated kinase 1A; DYRK1; DYRK1A; MNBEC 2.7.110EC 2.7.12; MNBH
Mass (kDA):
85.584 kDA

| Human | |
|---|---|
| Location: | 21q22.13 |
| Sequence: | 21; NC_000021.9 (37365573..37526358) |
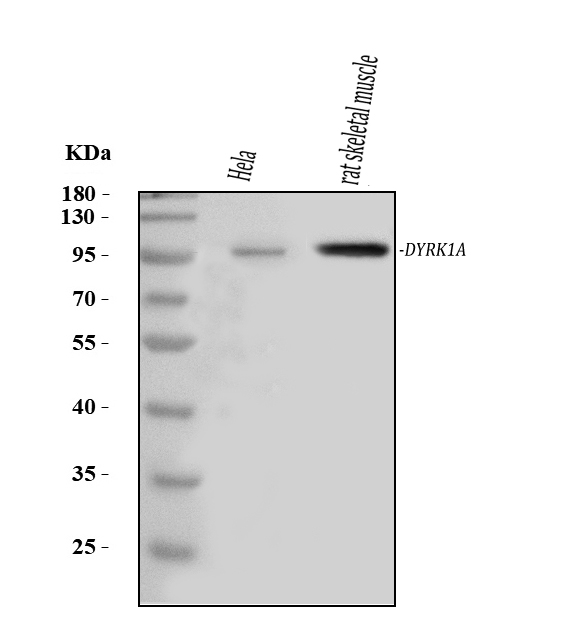
Ubiquitous. Highest levels in skeletal muscle, testis, fetal lung and fetal kidney.
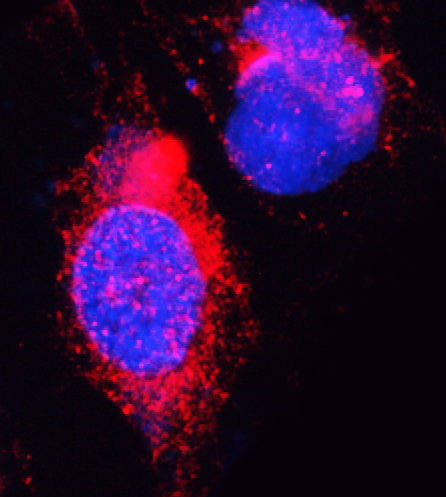
Nucleus. Nucleus speckle.





PMID: 8975710 by Song W.J., et al. Isolation of human and murine homologues of the Drosophila minibrain gene: human homologue maps to 21q22.2 in the Down syndrome 'critical region'.
PMID: 8872470 by Guimera J., et al. A human homologue of Drosophila minibrain (MNB) is expressed in the neuronal regions affected in Down syndrome and maps to the critical region.