This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
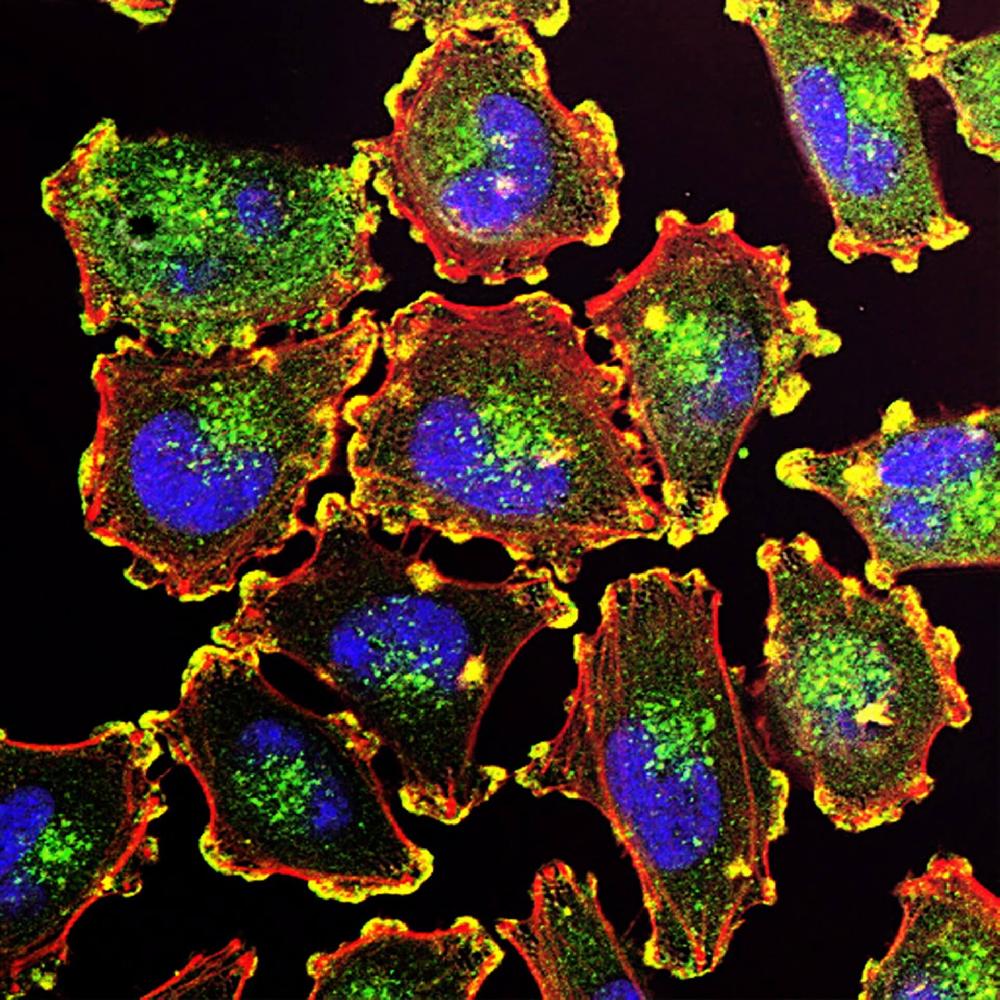



Facts about Serine/threonine-protein kinase ULK1.

Section of regulatory feedback loops in autophagy: acts both as a downstream effector and negative regulator of mammalian target of rapamycin complex 1 (mTORC1) via interaction with RPTOR. Activated via phosphorylation by AMPK and also acts as a regulator of AMPK by mediating phosphorylation of AMPK subunits PRKAA1, PRKAB2 and PRKAG1, resulting in negatively regulate AMPK activity.
| Human | |
|---|---|
| Gene Name: | ULK1 |
| Uniprot: | O75385 |
| Entrez: | 8408 |

| Belongs to: |
|---|
| protein kinase superfamily |

ATG1 autophagy related 1 homolog; ATG1; ATG1A; EC 2.7.11; EC 2.7.11.1; FLJ38455; FLJ46475; KIAA0722; serine/threonine-protein kinase ULK1; unc-51 (C. elegans)-like kinase 1; UNC51; Unc51.1; unc-51-like kinase 1 (C. elegans); Unc-51-like kinase 1
Mass (kDA):
112.631 kDA

| Human | |
|---|---|
| Location: | 12q24.33 |
| Sequence: | 12; NC_000012.12 (131894622..131923150) |
Ubiquitously expressed. Detected in the following adult tissues: skeletal muscle, heart, pancreas, brain, placenta, liver, kidney, and lung.
Cytoplasm, cytosol. Preautophagosomal structure. Under starvation conditions, is localized to puncate structures primarily representing the isolation membrane that sequesters a portion of the cytoplasm resulting in the formation of an autophagosome.





PMID: 9693035 by Kuroyanagi H., et al. Human ULK1, a novel serine/threonine kinase related to UNC-51 kinase of Caenorhabditis elegans: cDNA cloning, expression, and chromosomal assignment.
PMID: 11146101 by Okazaki N., et al. Interaction of the Unc-51-like kinase and microtubule-associated protein light chain 3 related proteins in the brain: possible role of vesicular transport in axonal elongation.











