This website uses cookies to ensure you get the best experience on our website.
- Table of Contents



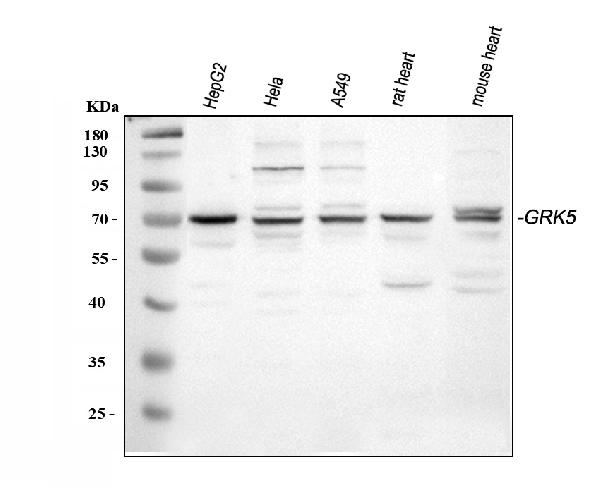
Facts about G protein-coupled receptor kinase 5.

Phosphorylates an assortment of GPCRs, such as adrenergic receptors, muscarinic acetylcholine receptors (more specifically Gi-coupled M2/M4 subtypes), dopamine receptors and opioid receptors. In addition to GPCRs, also phosphorylates various substrates: Hsc70- interacting protein/ST13, TP53/p53, HDAC5, and arrestin-1/ARRB1.
| Human | |
|---|---|
| Gene Name: | GRK5 |
| Uniprot: | P34947 |
| Entrez: | 2869 |

| Belongs to: |
|---|
| protein kinase superfamily |

EC 2.7.11; EC 2.7.11.16; FLJ39780; G protein-coupled receptor kinase 5; G protein-coupled receptor kinase GRK5; GPRK5; GPRK5FP2025; GRK5
Mass (kDA):
67.787 kDA

| Human | |
|---|---|
| Location: | 10q26.11 |
| Sequence: | 10; NC_000010.11 (119207571..119459745) |
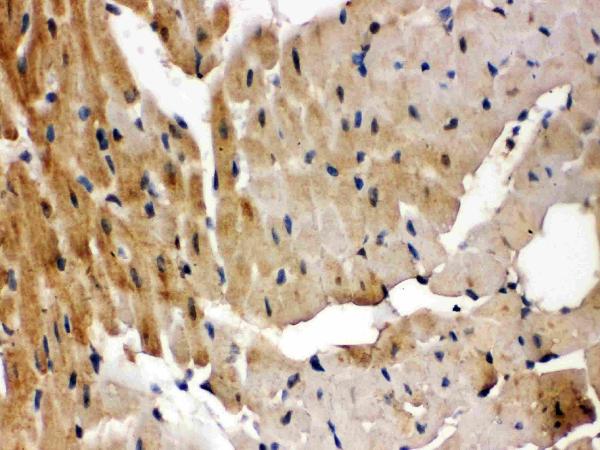
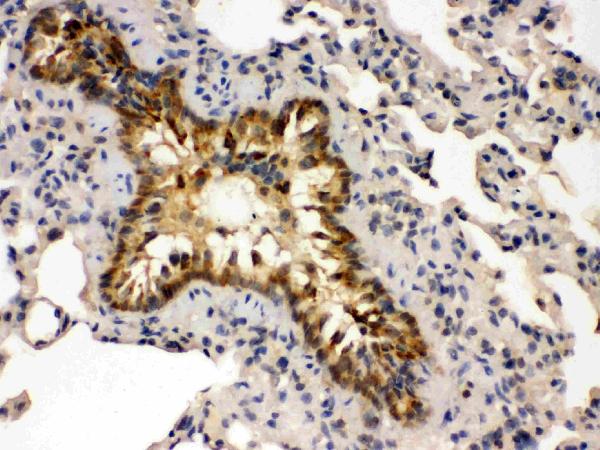
Highest levels in heart, placenta, lung > skeletal muscle > brain, liver, pancreas > kidney.
Cytoplasm. Nucleus. Cell membrane; Peripheral membrane protein. Predominantly localized at the plasma membrane; targeted to the cell surface through the interaction with phospholipids. Nucleus localization is regulated in a GPCR and Ca(2+)/calmodulin-dependent fashion.




GRK5 is a member the kinase-family and does not have post-translational modifications. The protein phosphorylates GPCRs as well as cytoplasmic and kinases but does not have any post-translational modifications. Its use in immunological and biochemical research is therefore limited. Continue reading to learn more about the GRK5 mark.
The electrostatic contour of the GRK5 kinase domain illustrates a bipartite surface charge distribution. The N-terminal is primarily negatively charged, while C-lobe tends to be positively charged. The plane of the protein houses the N-terminal membrane binding determinant. This bipartite surface-charge distribution could explain the kinase’s steric effect.
GRK5 has a monomer structure. The C-tail region extends from N-terminal membranebinding interface. Autophosphorylation of GRK5 occurs both intra and intermolecularly, and occurs at distinct sites along its NLT. Both types of autophosphorylation match the monomeric state GRK5 due to similar molecular weights.
Santa Cruz Biotechnology antibody sc-518005 detects GRK5 proteins that are overexpressed. It does not cross-react with GRK6 isoforms. Another antibody, CS-3982, recognizes GRK3-1 GRK3-1 protein sequences. It also shows a strong background band below the specific protein band. This property is not common for the GRK5 CS-3982 antibody.
The GRK5 protein was crystallized as a monomer in an asymmetric unit, while the GRK1 and GRK6 are dimers. This crystal structure does not exclude dynamic self-association under certain conditions. In-solution analysis confirmed that the GRK5 molecule is devoid of an in-solution dimer interface. These findings have implications on the biology of cancer cells.
GRK5 phosphorylates G-protein coupled receptors (GPCRs). In this study, we investigated whether GRK5 phosphorylates moesin, a type of integrin. Flag-moesin, an immunoglobulin, is phosphorylated at residue 666A. We also expressed Flag-moesin T66D in PC3 cells in a separate experiment. We also used an antimoesin antibody to purify GRK5 from kinase buffer. The final step was to test whether autoradiography detected the phosphorylated muesin protein.
The protein was first incubated for 1 hour on G-25 columns. The 0.2-mm GRK5 was incubated with nitrocellulose filters in binding buffer at 4 degC for 1 h. After this, a protein with 32P was added to the column. The mass-spectrometry analysis identified the bands. The authors concluded that phosphorylation of GRK5 affects focal adhesions and metastasis.
G-protein-coupled (G-protein -coupled) receptors are phosphorylated with GRK5. It has been suggested by GRK5 that a high level promotes G-protein -coupled receptor desensitization. Some G-proteincoupled receptors protect the heart and prevent further damage. Others promote a maladaptive phenotype. Additionally, b-ARs can contribute to pathogenesis by reducing inotropic reserve.
In addition to its role in regulating cell growth, GRK2 phosphorylates a subset of mTORC1 signal transducers and promotes activation of a tumor suppressor gene, p53. GRK2 phosphorylates ribosomal protein 2 and promotes translation activity for the 60S subunit. It interacts with IRS1 and promotes T cell activation by promoting cytokine release.
GRK2 is an important part of viral infection. It has been implicated as a catalyst in the uncoating and removal of flaviviruses like dengue virus or hepatitis C. It is also involved during the initial stages in influenza A virus infection. GRK2 is inhibited from replication by pharmacological inhibitors. Moreover, GRK2 siRNA kinase silencing limits viral replication in primary human airway epithelial cultures. GRK2 kinase activation is required for virus coating and is independent from beta-arrestin functions.
The structural organization in GRK5 shows two parts. The upper section is conical and the lower portion has two interconnected areas that resemble GRK5. A 2D average of 1100 particles reveals the GRK5 domains and the way it packs against Rho. Moreover, it also shows that phosphorylation of GRK5 is dependent on a phosphorylated Rho, whereas its non-specific activity is inhibited by its lack of interaction with Rho.
Proteins are built from a series of amino acids, typically 20 in number, and further tailored by covalent modifications that introduce new reactive groups and chemistries. These modifications control the activity and localization proteins. Although post-translational modifications (PTMs), are believed to occur on the amino acids side chains, these modifications can also affect the structure and function of proteins by controlling their backbone. This study highlights the importance PTMs play in controlling protein structure, and function.
A regulatory FDDY motif is formed by a large C-terminal extension from the kinase domain of GRK5. This region is near the RH Domain and is purple. This motif isn’t found in other GRKs. This motif has implications for binding GRK5 to adenine.
GRK5 dimer is known to interact well with phospholipids. GRK1 & GRK6 crystallize in dimers, but GRK5 is a single-mer. Sequence alignment revealed a unique Pro529 within the C-terminal area of GRK5. This Pro529 bends and flips the polypeptide chains back on themselves, changing the orientation of visible segments of the C-terminus. Similar to other GRKs, GRK5 also exhibits antiparallel C-terminal interface stabilization.
GRK5 is bipartite in its distribution of surface charges. The N-terminal region is predominantly positive charged, while C-lobes are negatively charged. The kinase domain consists of two hydrophilic determinants that are tightly coupled to each other. These interactions provide the structural foundation for allosteric effect. Boster Bio lacks a FDDY motifs in GRK5.
The structure and domain of GRK5 kinase are very similar to GRK6*sangivamycin's, PDB code #3NYN. The canonical ATP binding pocket contains the AMP-PNP ligand. Singivamycin, however, is found in the large cleft within the interlobe space. Both ligands can be found in the active area, with the AMP -PNP ligand occupying ATP-binding space. Both ligands contain conserved hydrogen bonds which serve to bind for the adenine bases. The AMP-PNP ligands have a hinge region that resembles the AST nucleotide interaction in PKB.
GRK5 -specific antibodies recognize a GRK5 signal containing a truncated mAb. They normalize the signal using the relative amounts of GRK forms. The antibodies were created with high affinity affinity and selectivity for the mAb that is directed against the human GRK5 wasoform. The antibodies detect GRK5 peptides but not GRK6.
The crystal structure for the GRK5 protein kinasedomain displays typical features, including a bilobalfold. The monomeric C-terminal portion is found in the kinase domain. The N-lobe consists of a canonical five-strand anti-parallel b-sheet while the C-lobe is situated under the interlobe hinge region.
The GRK5 proteins has a polybasic n and c terminus that targets the membranes. The polybasic regions are critical for membrane targeting, whereas other GRKs have distinct mechanisms. The N terminus lacks a lipid moiety while the C terminus far contains one. The C terminus also contains a pleckstrin-homology domain that binds acidsic lipids.
A pull-down assay was used to determine the binding between GRK5 & b2AR. The binding conditions were bicelles. In a pulldown assay 5 mM was solubilized into MNG-3 Micelles and immobilized onto M1 agarose. In this study, GRK5 showed a 60% increase in catalytic efficiency, while Km remained unaltered. This increase in kcat is attributed to enhanced binding to the receptor and increased phosphoryl transfer rate.
Mutagenesis (mutation) of the ionic locks residues further revealed structural differences between mutants and wild-type GRK5. Wild-type simulations showed transient separation of RH and kinase domains. However, mutations of alanine residues increased GRK5 dynamics and led to formation of two domains separated from each other by an ionic locking. These mutations caused the disruption of electrostatic interactions between the two domains, which led to enhanced phosphorylation for agonist-bound B2AR.
PMID: 7685906 by Kunapuli P., et al. Cloning and expression of GRK5: a member of the G protein-coupled receptor kinase family.
PMID: 8381058 by Ungerer M., et al. Altered expression of beta-adrenergic receptor kinase and beta 1- adrenergic receptors in the failing human heart.