This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
Murine immune system is comparable to that of humans, consisting of both innate and adaptive components, and its efficacy is regulated in cancer.
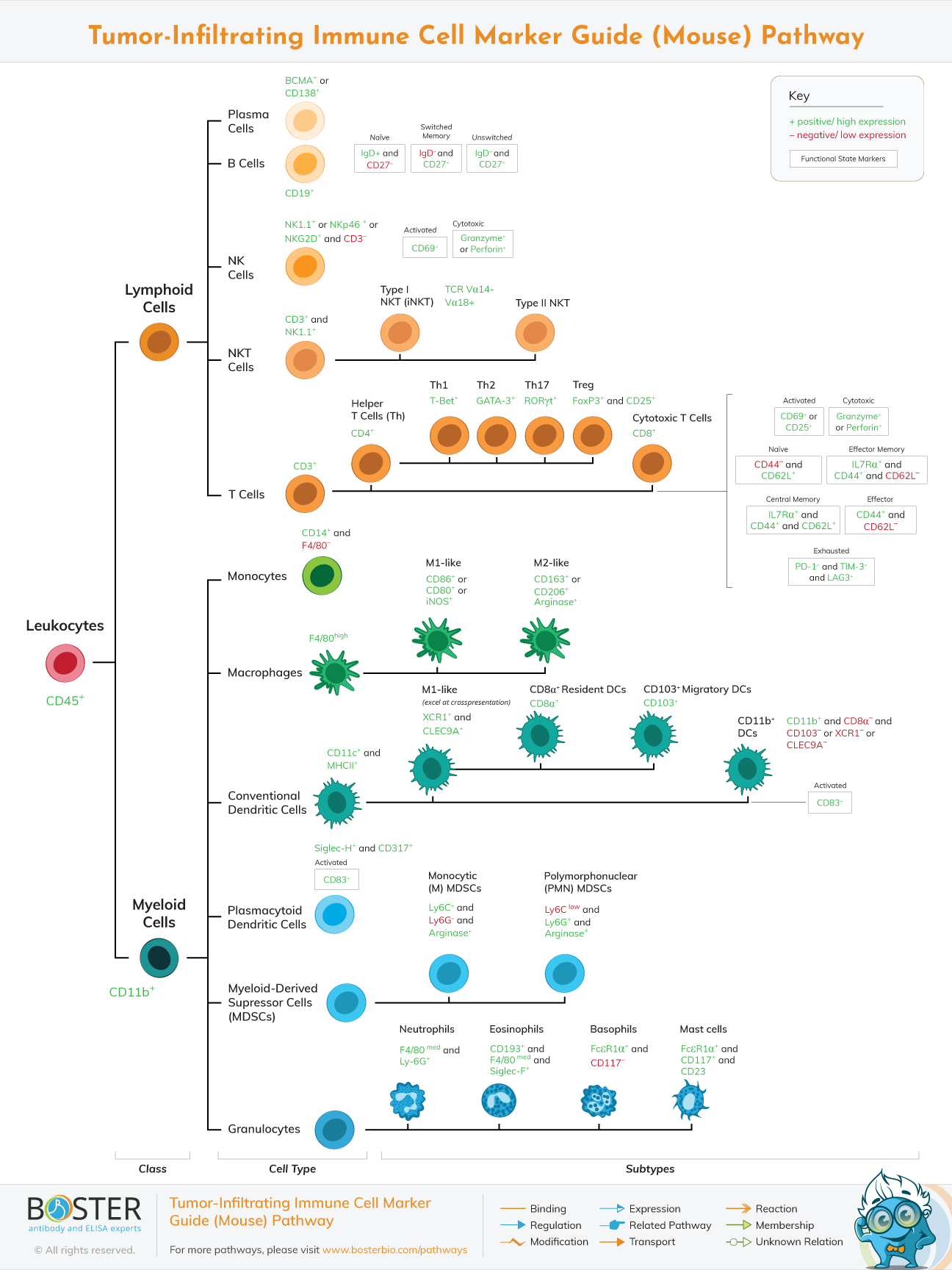
The mouse immune system is structurally similar to the human immune system, consisting of both innate and adaptive components, and its efficiency is regulated in cancer. Significantly, mouse models have advanced our understanding of cancer biology significantly, including the validation of cancer genes, the development of tumor biomarkers, and the evaluation of investigational therapies.
As is the case in humans, immune cells in mice regulate tumor growth and suppression through a complicated network of cytokines, chemokines, and growth factors. The many populations of mouse immune cells are described here.
CD3-expressing T lymphocytes in mice are classified as CD4+ helper or CD8+ cytotoxic. T cell activation increases CD69 and CD25 expression, which are widely utilized as activation markers. CD8+ cytotoxic cells lyse target cancer cells with serine proteases (granzyme) and pore-forming cytolytic proteins (perforin), whereas CD4+ helper cells coordinate the immune response via the secretion of diverse cytokines. As with human fatigued T cells, exhausted murine T cells exhibit a mixture of PD-1, TIM-3, and LAG-3; however, these molecules can also be produced following T cell activation. CD62L, IL7Ra, and CD44 expression can be used to identify naive, memory, and effector T cells in mice. The expression of transcription factors such as T-bet for antitumor Th1 cells and FoxP3/CD25 for protumor Treg cells distinguishes many cytokine-producing subsets of CD4+ cells.
Dendritic cells (DCs) from mice are antigen presenters for CD4+ and CD8+ T lymphocytes. Mouse cells, like human DCs, are classified into plasmacytoid and conventional subtypes. Plasmacytoid DCs co-express Siglec-H and CD317 and are capable of producing type I IFN, whereas typical DCs express CD11c and MHCII. DCs that excel in CD8+ T cell cross-presentation can be identified by their expression of XCR1 or CLEC9A.
As with humans, macrophages can eliminate apoptotic tumor cells in mice. Macrophages express an adhesion G-protein-coupled receptor known as F4/80. Macrophage polarization also occurs in mice, with M1-like cells identified by CD86, CD80, or iNOS expression and M2-like cells identified by CD163, CD206, or arginase expression. In mice, functional iNOS expression and IFN-induced mRNA induction are well documented.
Natural killer (NK) cells of the mouse innate immune system identify and kill cancer cells through a combination of activating and inhibitory receptors, which enables NK cells to eliminate cancer cells without causing injury to the host. They can be identified by the expression of NKG2D, NK1.1, or NKp46 in the absence of CD3. Additionally, NK cells release immunoregulatory cytokines. SHIP1, an SH-2-containing phosphatase, has been strongly associated with NK cell homeostasis and development in mice.
Finally, myeloid-derived suppressor cells (MDSCs) exhibit high quantities of CD11b, arginase, and GR1, which is composed of the membrane proteins Ly6C and Ly6G. MDSCs have been identified in cancers, bone marrow, blood, spleen, liver, and lung of mice.
They are classified generically into monocytic (CD11b+ Ly6G- Ly6Chi) and polymophonuclear (CD11b+ Ly6G+ Ly6Clo) populations, with the latter constituting the majority of malignancies. Distinguishing peripheral mononuclear cells from neutrophils, on the other hand, remains a challenge and is a subject of current research