This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
T cell receptors (TCR) are important for T cell activity and the establishment of the immunological synapse. It establishes a link between T cells and antigen-presenting cells (APCs).
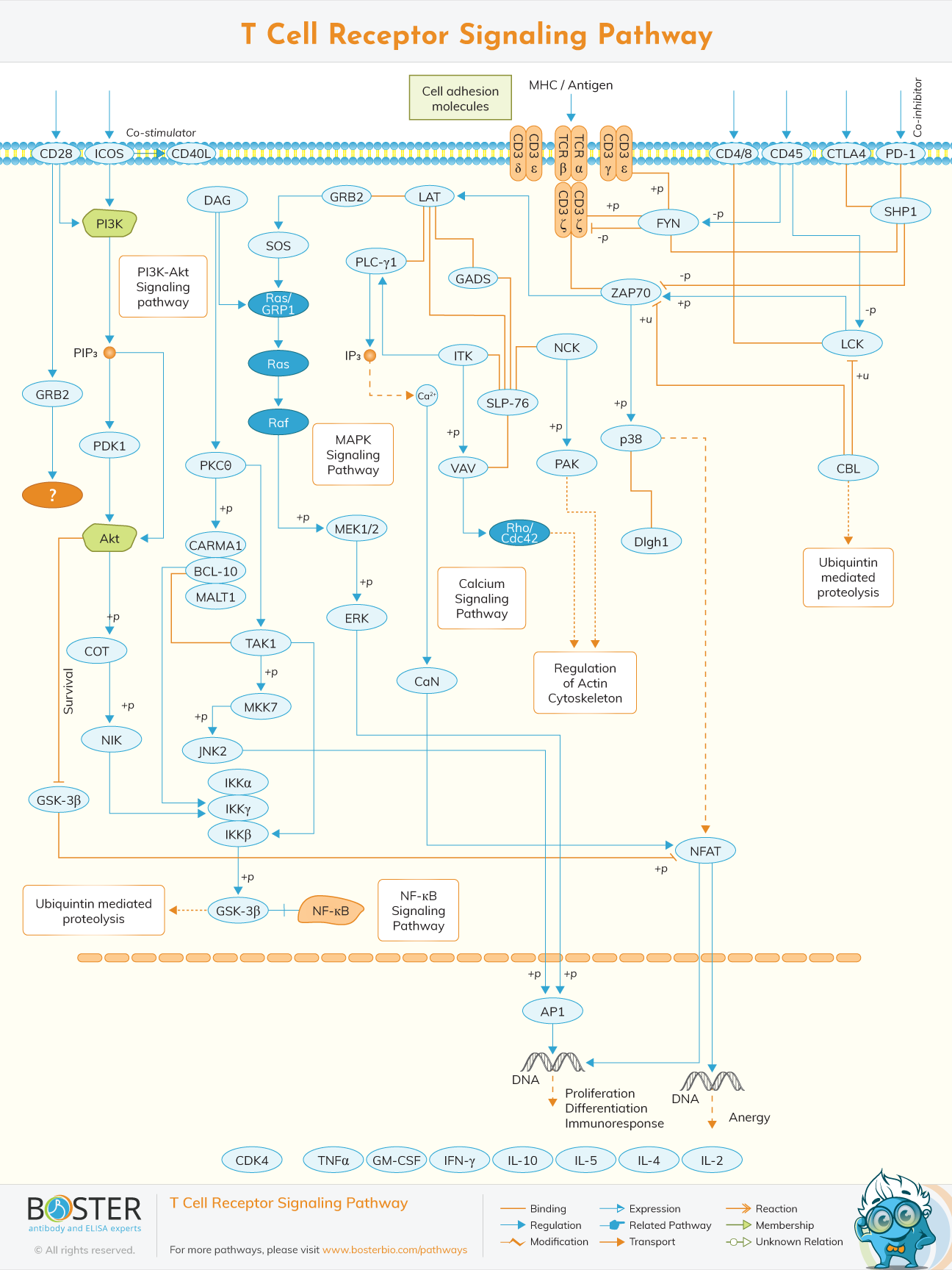
T cells are a type of lymphocyte that plays an important part in the immune system. T-cell receptor (TCR) is a complex of integral membrane proteins found on the surface of T cells that plays a role in T-cell activation in response to antigen. TCR activation is caused by MHC molecules on antigen presentation cells (APCs), which transmit antigen peptides to TCR complexes and initiate a series of cellular signaling cascades that lead to cellular proliferation, differentiation, cytokine release, and/or activation-induced cell death. TCR activation triggers a series of signaling cascades that control cytokine production, cell survival, proliferation, and differentiation, ultimately determining cell destiny. The phosphorylation of immunoreceptor tyrosine-based activation motifs (ITAMs) on the cytosolic side of the TCR/CD3 complex by lymphocyte protein tyrosine kinase is an early event in TCR activation (Lck).
Lck and other Src family tyrosine kinases are phosphorylated and activated by the CD45 receptor tyrosine phosphatase. The zeta-chain associated protein kinase (Zap-70) is recruited to the TCR/CD3 complex, where it gets active, increasing downstream adaptor or scaffold protein recruitment and phosphorylation.
The TCR heterodimer responsible for ligand recognition is made up of six distinct polypeptide chains. CD3 molecules (CD3, CD3, CD3, and CD3), when combined with the TCR heterodimer, have a unique tyrosine phosphorylation sequence motif known as immuno-receptor tyrosine-based activation motifs (ITAMs). TCR polypeptides have relatively short cytoplasmic tails, and CD3 molecules are in charge of mediating all proximal signaling events. The interaction between the TCR and the CD3 complex is critical for cell recognition events such as receptor construction, cell-surface expression, and signaling.
Several co-stimulatory receptors control TCR activation. TCR signaling is modulated by a variety of transmembrane receptors. TCR-induced proliferation and differentiation of naïve T lymphocytes are aided by CD28. CD28 supplies the T-cell with an initial adhesion ability by approaching the T-cell and APC membranes after interacting with B7-1 or B7-2. CD45 also modulates TCR signaling by altering the phosphorylation status of tyrosine kinases such as Lck and Fyn, as well as antagonizing the inhibitory effects of inhibitory proteins, promoting T-cell activation.
James P. Allison, a Nobel Laureate, was the first to discover the T-cell receptor in 1982. In 1984, Tak Wah Mak and Mark M. Davis discovered the human and mouse TCR cDNA clones, respectively. These discoveries revealed the identity and structure of the elusive TCR, dubbed the "Holy Grail of Immunology" in the past. This allows researchers from all over the world to study the TCR, resulting in significant research in the domains of CAR-T, cancer immunotherapy, and checkpoint inhibition
The Src family kinase Lck is activated by the interaction of MHC-II and CD4 or CD8, and proceeds to ITAMs on the,, and subunits of the TCR/CD3 complex upon activation of the TCR by antigen presented on MHC molecules. Phosphorylated ITAMs bind zeta-chain associated protein kinase (ZAP-70) to the TCR/CD3 complex, where it becomes activated, increasing downstream adaptor or scaffold protein recruitment and phosphorylation. ZAP-70's major substrates are the linker for activation of T lymphocytes (LAT) and the leukocyte protein SLP-76. The recruitment of a number of additional proteins involved in calcium mobilization, Ras pathway activation, and cytoskeletal rearrangement is triggered by phosphorylated LAT and SLP-76.
Phospholipase C1 (PL-C1) is a key protein that is recruited to tyrosine-phosphorylated LAT in response to TCR stimulation. By hydrolyzing phosphatidylinositol-4, 5-bisphosphate (PIP2) at the plasma membrane, activated PL-C1 produces the second messenger diacylglycerol (DAG) and inositol triphosphate (IP3). DAG activates a variety of proteins, including PKC isoforms and the MAPK/Erk pathways, both of which promote transcription factor NF-B activation, whereas IP3 binds to IP3 receptors (IP3Rs) on the surface of the endoplasmic reticulum (ER) and releases Ca2+. This event causes the Ca2+release activated Ca2+(CRAC) channels to open, enabling extracellular Ca2+ to enter. By breaking calmodulin's inhibitory functions, calcium-bound calmodulin (Ca2+/CaM) activates the protein phosphatase calcineurin. When calcineurin is activated, nuclear factor of activation T cell (NFAT) is dephosphorylated, allowing it to enter the nucleus and work with other transcription factors to enhance IL-2 gene production.
Ras is activated by the membrane recruitment of its exchange factors sos and RasGRP. Multiple members of the growth factor receptor-binding protein 2 (GRB2) family of adaptor proteins, such as GRB2, GRB2-related adaptor protein (GRAP), and GRB2-related adaptor protein-2 (GADS), bind tyrosine-phosphorylated LAT to facilitate the assembly of macromolecular signaling complexes needed for efficient T-Cell activation. The association of tyrosine-phosphorylated LAT with GRB2 allows GRB2 and GRAP-associated sos to be recruited to the plasma membrane and potentially activate Ras. Ras activation activates a number of serine/threonine kinases, including Raf1, MEK (MAPK/ERK Kinase), and dual-specificity kinases, all of which contribute to the activation of the mitogen activated protein kinases (MAPKs) superfamily. MAPKs activate kinases (and other signaling pathways) that activate the transcription factors NF-B and AP-1, which are required for the transcription of several critical genes involved in the T cell immunological response.
TCR signaling modifies the cytoskeletal architecture dramatically. T cell antigen recognition triggers a burst of actin polymerization at the immunological synapse, resulting in a lamellipodial sheet structure that extends throughout the APC's surface. The LAT–SH2-domain-containing SLP76 complex organizes two pathways that govern cytoskeleton changes after TCR triggering. One mechanism involves the adaptor–scaffold complex Src kinase-associated phosphoprotein of 55 kDa (SKAP55)–adhesion and degranulation promoting adaptor protein (ADAP), which regulates T-cell adhesion to the APC by increasing LFA1 integrin avidity. The other is dependent on VAV1, which binds to phosphorylated SLP76 via its Src-homology 2 (SH2) domain. VAV1 that has been phosphorylated on tyrosine promotes the Rho family GTPases, specifically Rac1 and Rac2. This activates a signaling cascade that regulates actin polymerization, TCR clustering, and the coalescence of glycolipid-rich membrane microdomains via the Wiskott-Aldrich syndrome protein (WASP) and the actin nucleating ARP2–ARP3 complex (GEMs). As a result, this route helps to organize the'signalosome' by concentrating receptors and signaling components.