This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
The B cell antigen receptor (BCR) is composed of membrane immunoglobulin (mIg) molecules and associated Igα/Igβ (CD79a/CD79b) heterodimers (α/β).
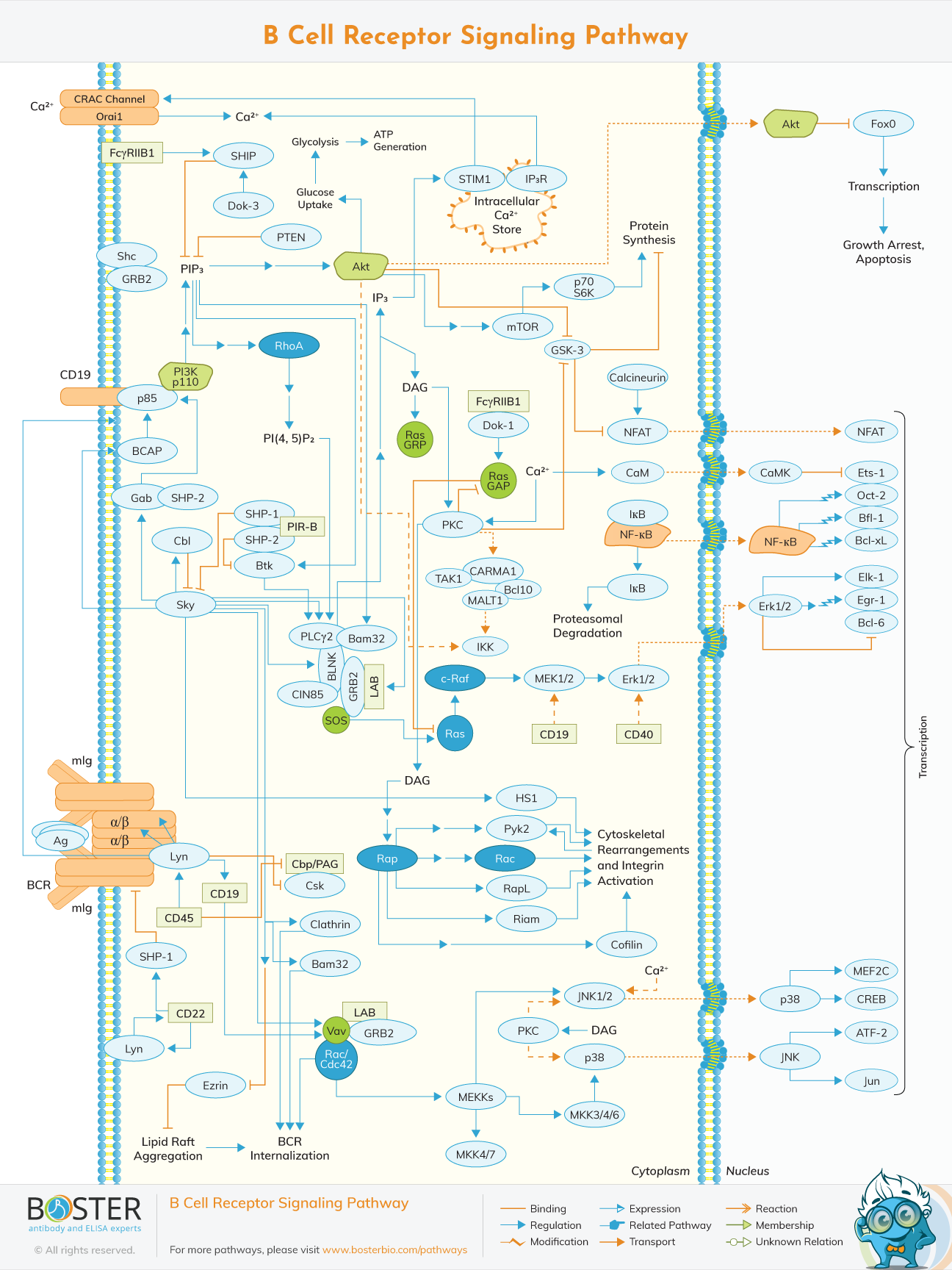
The antigen-binding transmembrane immunoglobulin (mIg), combined with the signal transduction moiety, Ig-/Ig-heterodimers(CD79a/CD79b), is a multiprotein complex produced on the surface of B lymphocytes. BCR signaling is required for appropriate B-cell growth, activation, proliferation, differentiation, and, as a result, humoral immune response.
Antigen engagement causes membrane movement and BCR component aggregation, which leads to phosphorylation of ITAMs in the cytoplasmic tails of CD79a and CD79b. The SRC family kinase LYN is responsible for the latter. The phosphorylated ITAMs attract the spleen tyrosine kinase (SYK) to the receptor, where it is activated by tyrosine phosphorylation and propagates signal activation to downstream signaling pathways. SYK activation is important for BCR signaling because it triggers the development of the BCR signalosome, adaptor proteins including CD19 and B-cell linker (BLNK), and Bruton's tyrosine kinases (BTK), as well as signaling enzymes like PLC2, PI3K, and Vav. Signals from those signalosomes activate and modulate downstream signaling pathways, such as the RAS/RAF/MEK/ERK pathway, which are important for B-cell fate decisions like proliferation, survival, differentiation, and cell death.
RAS can also activate ERK, which is triggered via the PLC2 pathway in B cells after BCR stimulation. RasGRP3, a guanine nucleotide–exchange factor, has recently been discovered to play a critical role in coupling the BCR to RAS activation via the PLC2/PKC pathway. RasGRP3 binds directly to RAF-1, the MAP3K in the ERK pathway, once activated. After that, activated RAF-1 phosphorylates MEK1/MEK2, which both activate ERK1/ERK2. Phosphorylated ERK1/ERK2 dimers form, which is required for ERKs to translocate into the nucleus and activate transcription of regulatory genes such as fos and jun, which regulate a group of genes involved in cell proliferation, survival, and differentiation.
LYN-dependent phosphorylation of CD19, which is found on the surface of B lymphocytes at all stages of their development, is required for PI3K activation. PIP3 is produced by activated PI3K and is involved in the transmission of signals downstream of PI3K. It causes membrane attachment and subsequent activation of numerous signaling proteins with pleckstrin homology (PH) domains. One of the most significant downstream effectors of PI3K, protein kinase AKT, can improve cell viability by inactivating proapoptotic proteins and transcription factors including BCL-2 and mTOR. AKT can also activate the expression of multiple anti-apoptotic genes via inducing the production of prosurvival genes and the degradation of IB, allowing NF-B to activate the expression of several anti-apoptotic genes. Furthermore, through inactivating GSK3, AKT is known to have a significant impact on the control of apoptosis and glucose metabolism. The operation of the PI3K-AKT signaling system is important for cell proliferation, growth, metabolism, and apoptosis in general.
Activation of PLC2 causes phospholipid hydrolysis, resulting in the second messengers inositol-1,4,5-triphosphate (IP3) and diacylglycerol (DAG), which drive intracellular calcium release and activate protein kinase C (PKC). The activation of nuclear factor of activation T cell (NFAT), a calcium-dependent transcription factor downstream of the calmodulin/calcineurin pathway involved in the formation of mature B cells, is then triggered by calcium-dependent signaling. Activated PKC, on the other hand, causes the activation of ERK, JNK, and p38. Furthermore, PKC activity causes the NF-B inhibitor (IB) to be phosphorylated and then degraded, activating NF-B, which plays a crucial role in B-cell activation, maturation, and survival.
A dynamic equilibrium between activating and inhibitory processes controls the volume and duration of BCR signaling. LYN, CD22 (the negative BCR co-receptor), and SH2 domain-containing tyrosine phosphatase-1 (SHP-1) are all limiting components in the negative control of BCR signaling. Transmembrane receptors like as FcRIIb1, CD19, and CD22, which connect with the BCR either constitutively or in a ligand-dependent manner, can alter signalosomes produced by the BCR. The immunoreceptor tyrosine-based inhibition motif (ITIM) in their cytoplasm, which is phosphorylated by LYN upon BCR stimulation, is responsible for their inhibitory action. Phosphorylated ITIMs recruit phosphatases such Src homology 2 (SH2) domain-containing phosphatase-1 (SHP-1) or SH2 domain-containing inositol phosphatase (SHIP), which stop BCR signalosomes from moving.
As a result, LYN controls both the positive and negative aspects of BCR signaling. ITIMs have recently been assigned a new immunoregulatory function: they can propagate inhibitory signals in particular configurations called inhibitory ITAMs (ITAMi).