This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
Histone methylation is required for proper developmental programming of the genome, and dysregulation of the methylation machinery can result in diseased states such as cancer. Indeed, cancer genome analyses identified lysine mutations in the H3K27 and H3K36 genes. These sites are more prevalent in certain types of cancer. As a result of the discovery of these enzymes, the impact modifications have on the genome, and disease-associated mutations, an entirely new therapeutic and biomarker space is emerging.
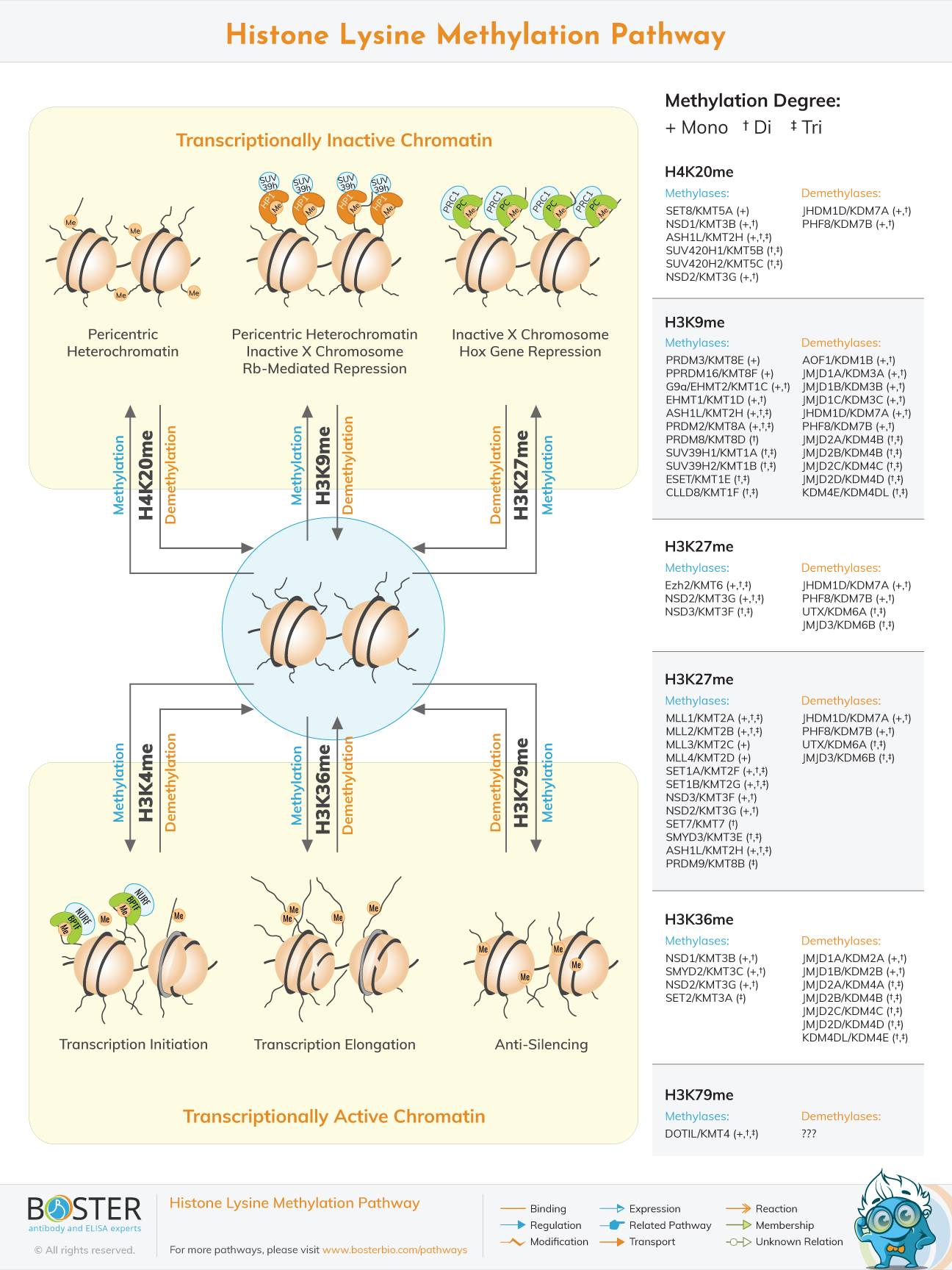
Histone modifications are critical epigenetic regulatory features that play a critical role in a variety of cellular processes. Lysine methylations are found at a variety of locations on histones' tail and globular domains, and their levels are precisely regulated by the action of methyltransferases ('writers') and demethylases ('erasers'). Additionally, distinct effector proteins ('readers') recognize specific methyl-lysines in a manner that is dependent on the amino acid sequence and methylation state of the neighboring amino acid. Histone lysine methylation dysregulation has been linked to a variety of cancers and developmental defects.
Histone methylation refers to the occurrence of N-terminal arginine or lysine in H3 and H4 histones. Methylation at the residue of the amino acid is mediated by histone methyltransferase. The function of histone methylation is mainly reflected in heterochromatin formation, gene imprinting, and X chromosome inactivation and in terms of transcriptional regulation, 24 histone methylation sites are currently found, of which 17 are in lysine and the other 7 are in arginine. Lysine may be monomethylated, dimethylated, and trimethylated.
Su(var) 3-9 is the first histone lysine methyltransferase discovered in Drosophila. It contains a conserved SET domain and is a plant ribulose diphosphate. In mammals, similar enzymes to Su(var)3-9 are SUV39H1 and SUV39H2, whereas in yeast, Su(var) 3-9 is similar to Clr4. These four enzymes catalyze only H3K9 methylation, whereas another methyltransferase G9a found in mammals can also catalyze H3K27 methylation. To identify additional methyltransferases, structural analysis methods are used to compare known protein structures. It is discovered that SET domain-associated proteins can be classified into four families: SET1, SET2, SUV39, and RIZ. Numerous them act as histone methyltransferases, in which various enzymes catalyze the methylation of various lysine sites. So far, dozens of lysine methyltransferases have been identified, classified into two major classes.
The first type of PRMT catalyzes the synthesis of methylarginine and asymmetric dimethylarginine, while the second type catalyzes the synthesis of monomethylarginine and symmetric dimethylarginine. PRMT1, PRMT3, RMT1/HMT1, PRMT4/CAMR1, and PRMT5 are members of the PRMT family. Only PRMT5 falls into the second category; the remainder fall into the first..
Histone H3K9, H3K27, H3K79, and H4K20 methylation acts as a transcriptional repressor. Methylation of H3K4 is a marker for transcriptional activation and is regulated by a variety of methyltransferases. The study discovered that histone H3K4 methylation expression is low in PC3 cells, whereas H3K9 methylation expression is high; PHI can increase histone H3K4 methylation while inhibiting H3K9 methylation. PHI has the ability to precisely control histone methylation expression. Transcription factors activate gene transcription, inhibiting PC3 tumor cell proliferation and inducing apoptosis. PHI has the ability to regulate the methylation status of H3K4 and H3K9 in the classical histone deacetylase inhibitor trichostatin, regardless of whether PHI is a histone methyltransferase or acts via other pathways. Histone methyltransferase activation or recruitment alters the methylation status of H3K9 and H3K4, but the precise mechanism remains unknown. Inhibitors of histone methyltransferase (HMTis): HMTis have been shown to specifically target histone lysine methyltransferases (HKMTs), including G9a, GLP1, and DOT1L. BIX01294 was the first inhibitor of HKMT to be discovered. Although BIX01294 binds to G9a and GLP1, its low affinity and cytotoxicity preclude it from being used. E27 and UNC321 are inhibitors of generation 2 whose chemical structure contains a 7-alkoxy amino group, which increases their affinity for the enzyme. Additionally, UNC638 contains a 7-alkoxyamine group, which promotes cell efficiency, reduces toxicity, and broadens application possibilities.
Other small molecule inhibitors, such as AZ505, inhibit the oncoprotein SMYD2 expression efficiently and specifically by binding to the substrate binding site of HKMTs. EPZ004777 is a SAM analog that competes for binding with DOT1L and inhibits DOT1L. The fungal metabolites chitin and carprofen are examples of compounds that bind to SAM. Along with HKMTis, potent inhibitors of protein arginine methyltransferases (PRMTs) have been discovered, including AzaAdoMet3, an amine analog of adenosylmethionine, and purine or pyrazole inhibitors.