This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
Microtubules are essential for cell polarity, polarized cell motility, intracellular vesicle trafficking, and chromosomal segregation during mitosis. Microtubules (MTs) are nonequilibrium polymers of /-tubulin heterodimers that undergo GTP hydrolysis on the -tubulin subunit after construction.
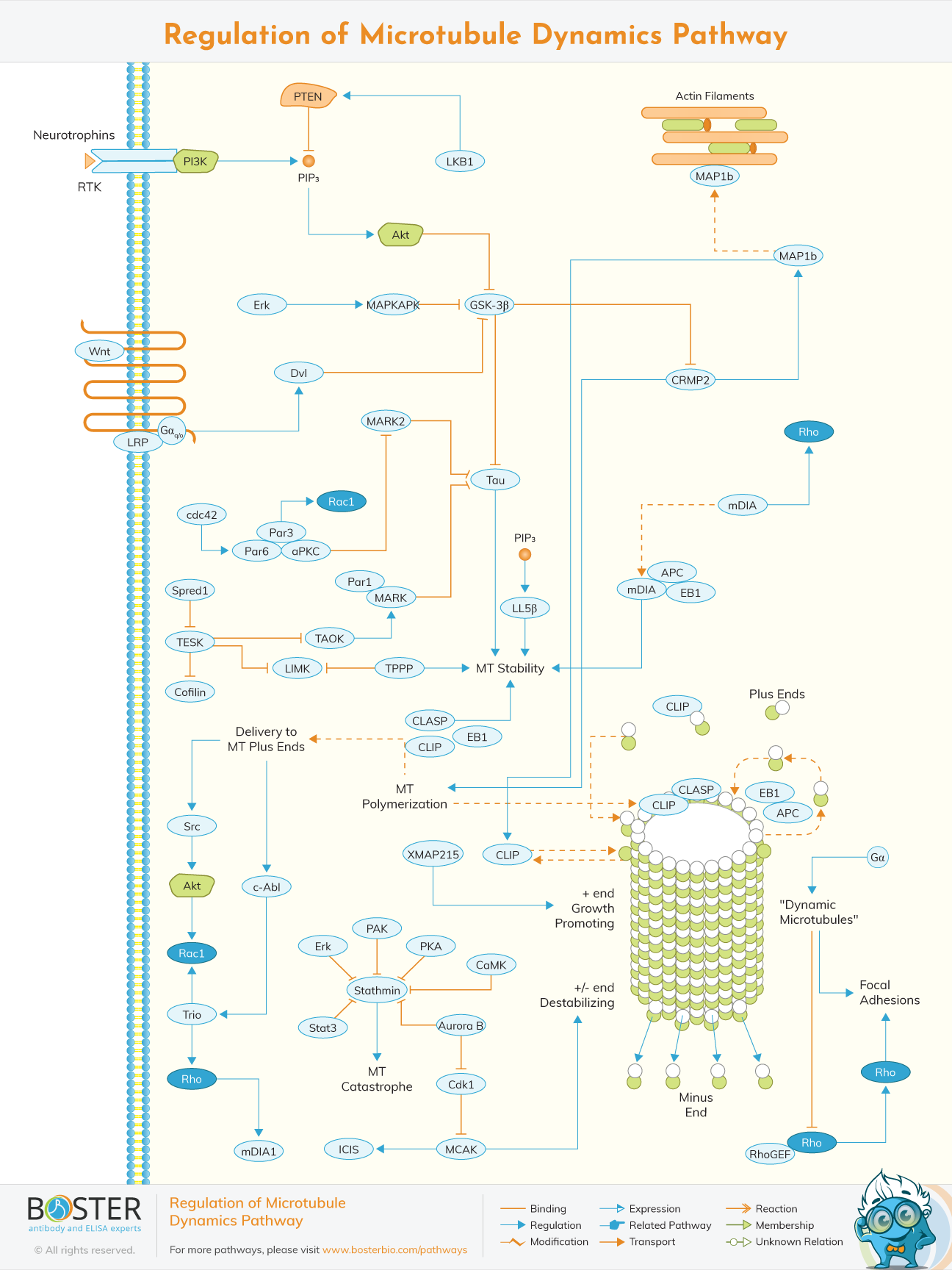
Microtubules are the major components of cytoskeleton.they are found in the eukaryotic cells are are involved in mitosis,maintenance of cell shape and intracellular transport. They are formed by the polymerization of a dimer of two globular proteins alpha and beta tubulin into protofilaments that can then be associated laterally to form a hollow tube. Due to their participation in most cellular functions,they must be regulated in response to extracellular and intracellular signals.because of complexity and numerosity of microtubule dynamics,the connection and regulation of microtubule dynamics remains indistinct.
The majority of microtubules are formed from organizing centers. The most common microtubule activity is dynamic instability, which is characterized by sluggish plus end expansion followed by rapid depolymerization (“catastrophe”) and subsequent rescue. Although the minus ends of microtubules exhibit dynamic instability, albeit at a slower pace than the plus ends, the minus ends are often capped and attached to MT organizing centers, and so do not frequently engage in microtubule dynamics.
The discovery of the microtubule -mediated processes like cell locomotion is traced back in the 17th century and in particular 1677 when Leeuwenhoek observed them via the microscopes.The discovery of fibrous nature of the flagella was later discovered through the improved light microscope and in 20th century,confirmed with the electron microscope.
The balance between dynamically unstable and stable microtubules is mostly controlled by proteins that bind either tubulin dimers or assembled microtubules. Stathmin, which sequesters tubulin and improves MT dynamics by increasing catastrophe frequency, and collapsin response mediator protein (CRMP2), which promotes tubulin dimer addition to microtubule plus ends, are two proteins that bind tubulin dimers. Other proteins that interact with assembled MTs include those that bundle MTs (e.g., MAP1c), those that stabilize MTs (e.g., tau), and those that keep MTs dynamic (MAP1b). GSK-3, a kinase that is normally active under baseline growth circumstances but becomes locally inactive in response to cues that promote MT growth and dynamics, is a key signaling route that governs MT dynamics.
Tubulin undergoes a variety of post-translational changes, including acetylation, polyglutamylation, and poly-glycylation, which have been found to modify its interaction with certain MT motors as well as other proteins that can impact MT stability and dynamics.
In addition to the aforementioned variables, numerous MT motor proteins, as well as non-motor proteins, contribute to the dynamics of MTs. Proteins like Xenopus microtubule associated protein 215 (XMAP215) enhance MT formation by binding to tubulin dimers and allowing them to be incorporated into the developing plus end. XMAP215 may also compete with some of the MT plus end binding proteins (+TIPS), among which EB1 appears to be the main organizer. Complexes formed by the adenomateous polyposis coli (APC) protein and plus end binding proteins help to stabilize MTs by lengthening the MT elongation phase. Several nonmotile kinesins from the kinesin-13 family induce MT instability. In vitro, the mitotic centromere associated kinesin (MCAK), one of the best researched kinesin-13 family members, binds both plus and minus MT ends. By decreasing the lateral contacts between the protofilaments, the binding of MCAK to an MT end is considered to speed the transition to disaster.