This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
Microtubules are essential for cell polarity, polarized cell motility, intracellular vesicle trafficking, and chromosomal segregation during mitosis. Microtubules (MTs) are nonequilibrium polymers of /-tubulin heterodimers that undergo GTP hydrolysis on the -tubulin subunit after construction.
Actin filaments are formed in the eukaryotic cells and are essential for many of the movements especially those involving the cell surface without action filaments e.g an animal cell could not crawl along a surface , engulf a large particle by phagocytosis or divide into two. Actin is unstable but it can be stable structures in cells e.g the contractile apparatus of muscles.
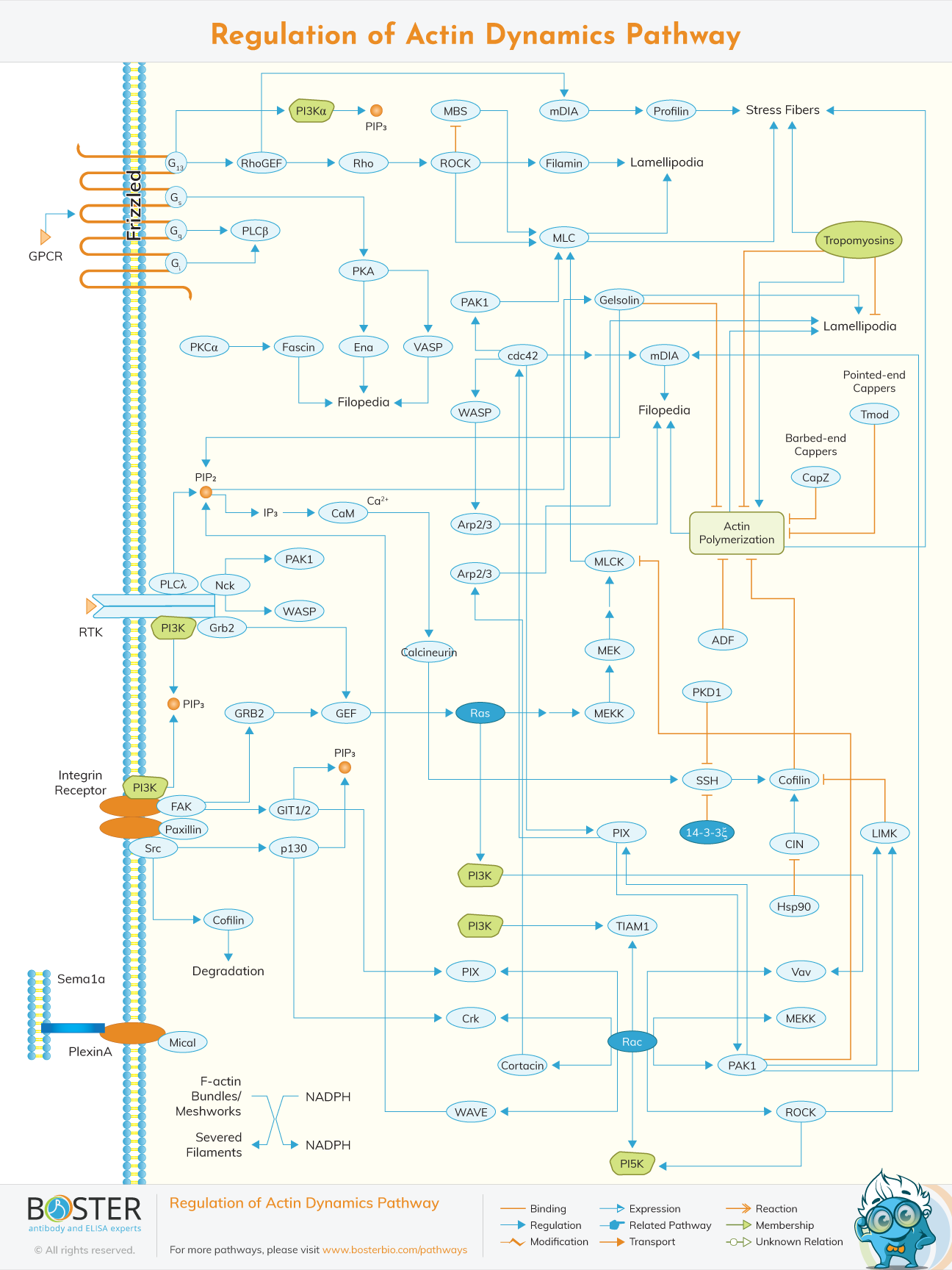
Actin dymanics pathway can be regulated in various number of ways including the phosphorylation/dephosphorylation of the colifin, through the Rho family GTPases, G-actin binding proteins, the Arp2/3 complex among others.
Actin was first observed experimentally in 1887 by W. D. Halliburton, who extracted a protein from muscle that "coagulated" preparations of myosin and which he dubbed "myosin-ferment" (Halliburton 1887). However, Halliburton was unable to further characterize his findings and the discovery of actin is generally credited instead to Brúnó F. Straub.
Signals received by cell receptors through chemical messengers like the growth factor, hormones and cytokines are transmitted to Rho family GTPases and more so Rho, Rac, Cdc42. Actomyosin, movement, lamellipodia and filopodia formation develops due to the dispatchment of the intergraded signalling pathways to the cytoskeleton effector proteins.
Cell migration, cytokinesis, endocytosis, morphogenesis during development can be impaired due to the ectopic expression of dominant negative mutants of Rho family GTPases in the mammmalian cells.Cdc42/Rac interactive binding motif is a short number extension approximate 18 amino acIds found in the bigger number of proteins that associates with Cdc42 and Rac. Wasp, formins and IRSp53 have been characterized as potential Rho family effector proteins after a database search for CRIB motif containing proteins. These proteins contain multiple domains like FH2,WH2 and the actin-binding domains.they are also typically self-inhibited by internal interaction which are commonly released by the binding of the GTPases thus achieving two major functions; activation and recruitment of specific loci at the membrane.
In the resting state,the c terminus WCA domains of WASP is masked by intramolecular interaction with the CRIB motif located towards the N-terminus of WASP. Auto-inhibited conformation is the term used to refer this conformation. When stimulated,WASP moves to the plasma membrane where it binds simultaneously to GTP-Cdc 42 via its CRIB domain and to the plasma membrane via the basic domain located adjacent to the Crib domain.
The activated conformation of WASP referred to as the open conformation releases the WCA inhibition , paving way for the recruitment and activation via the Arp2/3 complex.Formins are believed to be activated in a similar manner.
Profilin, ADF/Cofilin and WH2-related protein Thymosin-beta-4 are some members of this group.they play a crucial role in controlling the pool of unpolymerised Actin monomers and there by regulating the pointed and barbed end addition/dissociation of Actin monomers.most structures of complexes of actin with ABPs have been isolated.
Most of these proteins have a conserved amphipathic alpha helix binding in the hydrophobic or target binding cleft in sub domain 1 and 3 and the barbed end of the Actin monomer. Missing in metastasis (MIM), WASP-family proteins and ciboulot are examples of WH2 domain.they share common features in that they has an N terminus alpha helix followed by an extended region.
The hydrophobic cleft in the action monomer faces the N -terminal alpha helix. C-terminal portions of the WH2 domains of different proteins follow a similar path on the action monomer,streaming towards the pointed end of the action monomer I.e towards the subdomains 2 and 4.
Colitin phosphorylation reduces filament breakdown by abolishing its action binding activity.colitin is also regulated by the Rho GTPases and the Albeit in a less direct way.
Therefore cofilin is regulated by Rho via Rho kinase that phosphorylates LIM kinase which inturn phosphorylates cofilin.
Through this signaling cascades, action assembly in many cells types is modulated in response to various extracellular stimuli.
Nucleation and elongation factor regulating the de novo formation of actin filament in cells is been worked on lately.Arp2/3 nucleaters including formins,spire,cobl,Vopl,VOpF and Lmod have been discovered following the Arp2/3 discovery.
Apart from the formins, the rest use the WH2 domain for interaction with actin. The formation of small oligomers of 2-4 actin subunits is kinetically unfavourable .Furthermore,actin monomers frequently associate with profilin and T beta 4 which control their incorporation into filaments .in conjunction ,they limit the spontaneous nucleation of actin filaments in cells thereby creating an opportunity for cells to actively regulate the de novo polymerisation of actin by using actin filament nucleation and elongation factors.
When the Arp2/3 complex is inactive,the Arp are separated and their nucleotide binding clefts are wide open .It is suggested that when being activated, the Arps adopt an actin filament-like conformation.
These are WH2-based filament elongation factors and are involved in axon guidance and migration of cancer cells. They form tetramers by their c-terminus called coiled coil domain and thought to bind to the barbed ends of actin filament bundles catalyzing their synchronized elongation against the plasma membrane.so, the filament bundles grow to form cellular structures like filopodia used by cells for irritability.
They are modular, containing N-terminal Ena/VASP homology 1,central pro-rich and c-terminal EVH2 domains. Evh1 domain binds to the consensus sequence motif FPPPP to their sites of action.EVH2 is divided into G and F-actin binding domains and C- terminus coiled coil tetramerisation domain regulatory proteins and profilin actin binds to the pre-rich region.these regions are commonly found in the formins and NPFs of Arp2/3.