This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
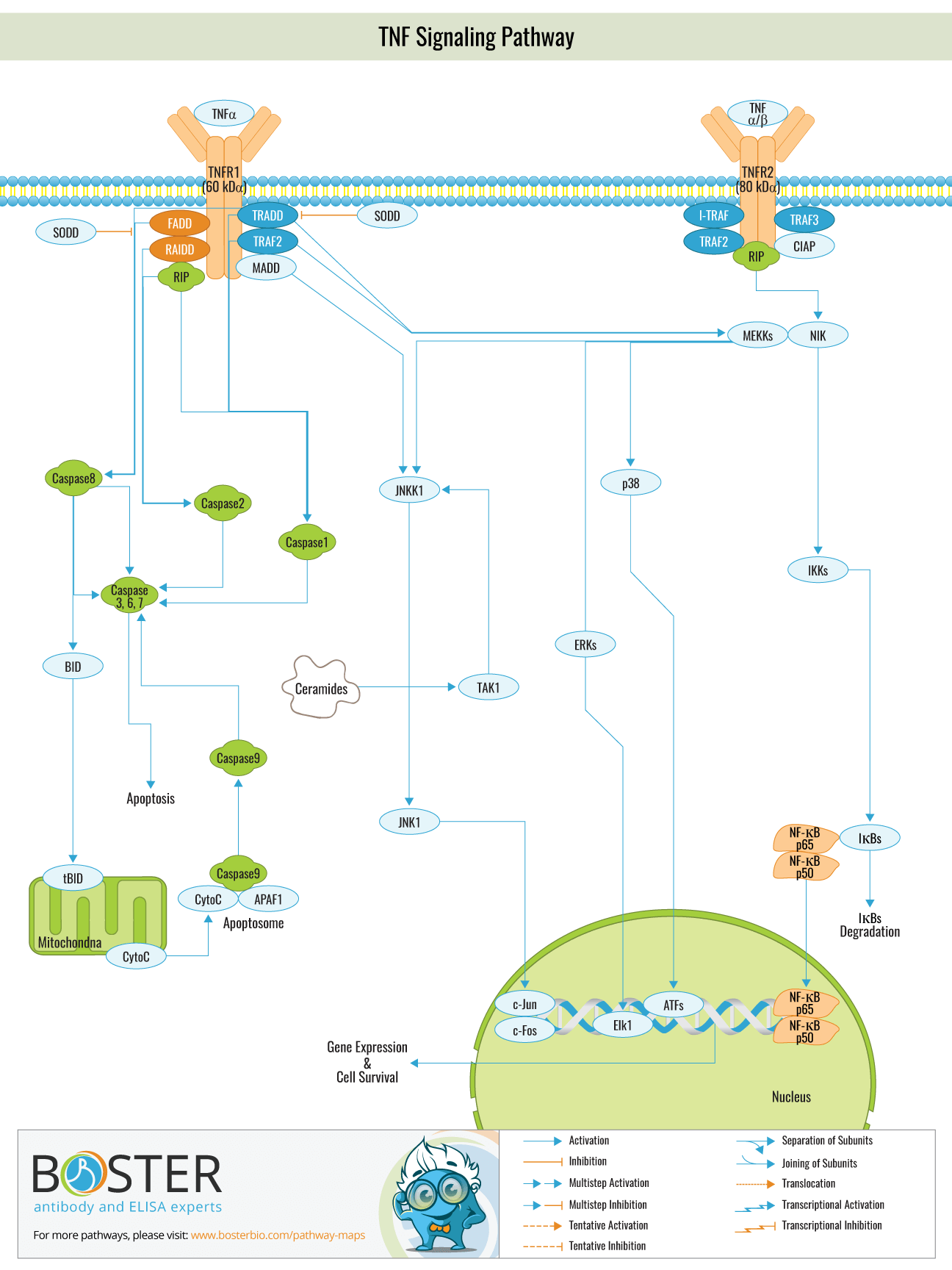
Tumor necrosis factor (TNF) is a type of cytokine that exerts a variety of biological functions. By binding to specific receptors on the cell membrane, it promotes cell growth, differentiation, apoptosis, and inflammation. TNF- is a member of the TNF family that has the ability to activate ERK (extracellular signal 2 regulated protein kinase), Caspase protease, and JNK. Additionally, it has independent pathways for cytotoxicity, antiviral activity, immune regulation, and apoptosis. Due to the fact that TNF- is intimately linked to cell homeostasis and a variety of human diseases, including tumors, research on the TNF- signaling pathway has become a hot topic in biomedical research over the last decade.
The primary role of TNF signaling pathway is in the regulation of immune cells. TNF, being an endogenous pyrogen, is able to induce fever, apoptotic cell death, cachexia, inflammation and to inhibit tumorigenesis and viral replication and respond to sepsis via IL1 & IL6 producing cells.
TNF- alpha is primarily secreted by activated mononuclear macrophages and is classified as a type II membrane protein that exists in trimers. TNF-alpha is available in two forms: 17 kD secreted soluble sTNF- and 26 kD membrane-associated mTNF-alpha . mTNF-alpha is widely believed to be a precursor to sTNF-, which is cleaved from the membrane by membrane metalloproteinase and shed as STNF-. TNF- performs its biological function by binding to the cell surface TNF-R receptor. TNR is classified into two subtypes: TNF-Rl (55 kD) and TNF-R2 (75 kD), both of which are type I membrane proteins composed of four components: a signal peptide, an extracellular domain, a transmembrane domain, and an intracellular domain. Both proteins have a low degree of homology in their intracellular domains. TNF-R1 contains a death domain (DD), whereas TNF-R2 does not, implying that the two receptor types engage distinct signaling pathways and mediate distinct biological effects. T
NF-R2 is more susceptible to mTNF activation than sTNF, and it is speculated that mTNF plays a significant role in ligand delivery in sTNF-mediated cellular responses, ie, TNF-R2 binds to TNF first, then delivers TNF to TNF-R1 and increases the affinity of TNF-R1 and TNF. This partially explains how low concentrations of TNF can initiate biological reactions.
TNF mediates two signaling pathways in vivo and one signaling pathway for apoptosis; following TRADD(TNF-R1 associated death domain) recruitment of TNF-R1, TNF-R1 physically recruits FADD to the TNF-R1 complex. The N-terminus of FADD contains a novel domain known as the MORT domain or death effector domain (DED), which activates Caspase 8, a member of the ICE/CED3 cysteine protease family. Activated Caspase-8 then activates other ICE/CED3 proteases. Thus, a cascade amplification reaction involving caspasse-8 protease induces apoptosis. TNF- stimulates the cell to form the TNF-R1 complex, which activates via a series of protein phosphorylation events. MAPKKK then activates MAPKK, which activates MAPK, which activates the JNK signaling pathway. JNK and P38 are both members of the MAPK family. The study discovered that the majority of the upstream molecules that activate JNK are members of the MAPKK family, specifically MKK4 and MKK7. MK is more likely to phosphorylate JNK Tyr85 than MKK7, although activation of JNK requires phosphorylation at both sites. P38 is primarily composed of MKK3.6 and MKK, both of which are MAPKK family members.
The study discovered that knocking out the makk1 gene in mice inhibited JNK activation as well, indicating that MEKK1 is required for the JNK signaling pathway to function properly. A subsequent study revealed that MEKK1 was activated in response to TNF stimulation, and when combined with TRAF2, phosphorylated MKK4, thereby activating JNK. However, it is unknown whether other kinases exist between MEKK1 and TRAF2.
NF-KB inhibits TNF-induced apoptosis, and activation of NF-KB has the primary cellular effect of promoting the expression of inflammatory proteins and inhibiting apoptosis. It exerts anti-apoptotic activity primarily by increasing the expression of apoptotic inhibitors. c-IAP, c-FLIP, and TRAF2 are all apoptotic inhibitors. C-IAP can inhibit caspase-3 or caspase-7 activity directly, as well as the activation of caspase-6 or caspase-9 precursors, thereby inhibiting TNF-induced apoptosis. A20 exerts a negative regulatory effect on the NF- signaling pathway: mice lacking the a20 gene were unable to reduce TNF- regulation, resulting in chronic inflammation and cell death, indicating that A20 was an NF- inhibitor.
A20 is a deubiquitination enzyme with an N-terminal domain that can deubiquitinate the lys63-linked ubiquitin chain in the RIP molecule and a C-terminal domain that contains seven C2/C2 zinc finger domains that act as a ubiquitin ligase and can connect the ubiquitin chain at lys48 in RIP, allowing it to enter the protease complex degradation pathway.