This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
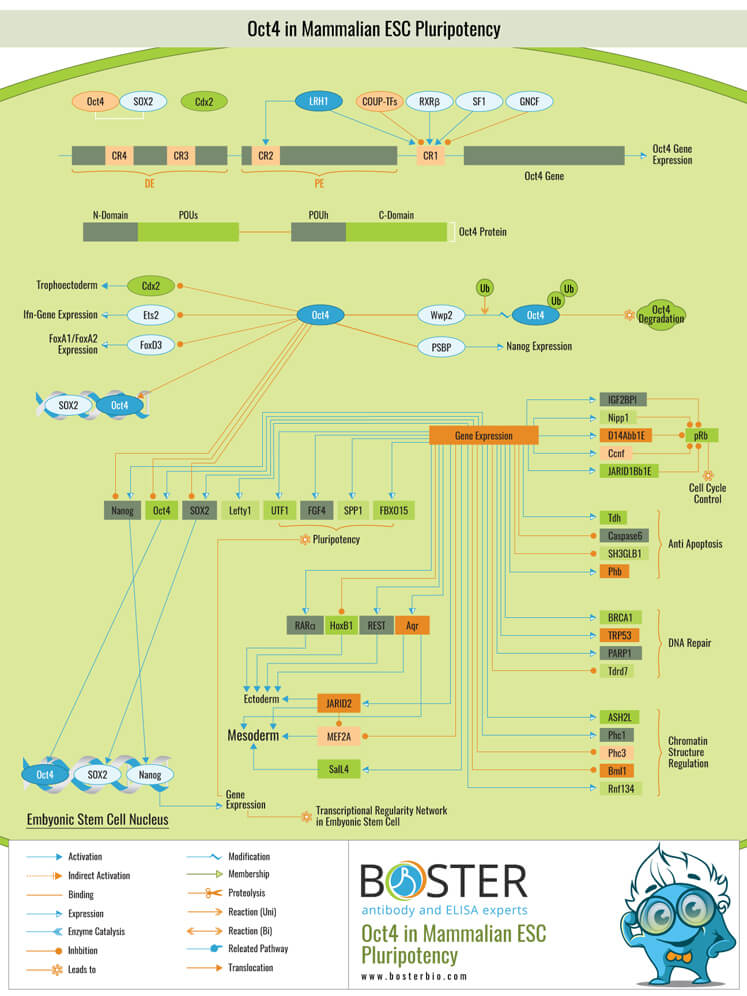
Stem cells are undifferentiated cells that can differentiate into virtually any type of cell in our body. They are defined by their capacity for self-renewal and maintenance of pluripotency. ESCs (Embryonic Stem Cells) must tightly regulate their differentiation status in order to achieve the desired developmental outcome. Hundreds of genes, including several transcription factors, have been identified whose expression patterns are highly correlated with ESC differentiation. A relatively small number of transcription factors are required to control the development of an entire organism. These transcription factors organize the correct temporal–spatial expression of developmental genes by forming multiprotein complexes on DNA.
The process results in the formation of functional partnerships, with the combination of each factor's activity eliciting specific transcriptional outcomes rather than its individual activity. Oct4 (Octamer Binding Transcription Factor-4) and Nanog have been identified as critical transcription factors for the maintenance of the pluripotent state of ESCs
Oct4, also known as Oct3, is a POU (Pit-Oct-Unc) family homeodomain transcription factor. The POU family of transcription factors can activate their target genes' expression by binding to an octameric sequence motif of the AGTCAAAT consensus sequence. Oct4 (POU5F1) is required for self-renewal, pluripotency, and lineage commitment in ESCs, embryonic epiblasts, and pluripotent stem cells (Primordial Germ Cells). Oct4 expression levels should be tightly regulated in order to control cell fate. Oct4 expression changes facilitate differentiation and result in the specification of Ectodermal, Endodermal, or Mesodermal primitive progenitors. Oct4 protein is composed of three domains: N-terminal, POU, and C-terminal. The POU domain is composed of two structurally distinct subdomains: a 75 amino acid carboxyl-terminal POU specific (POUs) region and a 60 amino acid carboxyl-terminal homeodomain (POUh). Both domains are connected by a variable linker of 15 to 56 amino acids and make specific contact with DNA via a helix-turn helix structure. Outside of the POU domain, regions are not required for DNA binding and have little sequence conservation. Proline and acidic residues are abundant in the N-terminal domain (N-domain), whereas Proline, Serine, and Threonine residues are abundant in the C-terminal domain (C-domain). The N-domain is required for transactivation.
Additionally, the C-domain is involved in transactivation. The Oct4 C-domain activity is cell type specific and is regulated by phosphorylation, whereas the N-domain activity is not. The Oct4 POU-domain is unique in that it serves as a site of interaction for cell type-specific regulatory factors
The precise mechanisms regulating the activation and expression of Oct4 genes remain unknown. Two regulatory elements, a Distal and a Proximal Enhancer, have been identified as stem cell-specific enhancers of the Oct3/4 gene, to which a large number of positive and negative regulators are recruited. There are four highly conserved regions in the Oct4 promoter/enhancer elements of Human, Bovine, and Mouse, designated as CR1 (Conserved Region-1), CR2, CR3, and CR4, respectively. CR1 is located immediately upstream of Exon 1 and downstream of Proximal Enhancer. Each enhancer contains multiple transcription factor binding sites that can either activate or inhibit Oct4 expression. Furthermore, methylation of these regions inhibits the expression of Oct4 in differentiated cells. Numerous positive and negative regulators regulate the Oct4 gene's expression. Among them, it is known that members of the orphan nuclear receptor superfamily that can bind to the Proximal Enhancer regulate Oct3/4 expression. LRH-1 (Liver Receptor Homolog-1), also known as NR5A2, is a putative positive regulator of the Oct3/4 transcription factor. SF1 (Steroidogenic Factor-1) and RXR-Beta are two additional positive regulators (Retinoid X Receptor-Beta). In comparison, GCNF (Germ Cell Nuclear Factor) or NR6A1 may act as a negative regulator of Oct3/4. COUF-TFI/II (Chicken Ovalbumin Upstream Promoter-Transcription Factors-I/II), which are encoded by NR2F1 and NR2F2, respectively, act as negative regulators of Oct3/4 expression. The balance of these positive and negative regulators may be critical for determining the precise level of Oct3/4 expression in response to extracellular stimuli. Additionally, Oct4 can be degraded via interaction with a murine E3 Ub ligaseWwp2 (WW domain containing E3 Ubiquitin Protein ligase-2). Wwp2 can interact directly with Oct4 via WW domains. It promotes Oct4 ubiquitination in vitro and in vivo.
Once activated, the Oct4 protein can function as an activator or repressor of target genes, thereby preserving stem cell pluripotency. Oct4 typically acts in concert with other regulators to promote the expression of specific target genes in defined cell types and developmental stages. SOX2 (SRY (Sex Determining Region-Y) Box-2) and Nanog are the most frequently used regulators that cooperate with Oct4 to activate Oct3/4 target genes. SOX2 is a transcription factor with an HMG domain that binds to the consensus motif CATTGTT. Oct4 and SOX2 interact with thousands of regulatory sites throughout the ESC genome. They reciprocally regulate the transcription of POU5F1 and SOX2 in ESCs via the Oct4-SOX2 complex. Additionally, Oct4 and SOX2 positively regulate Nanog, indicating that ESCs are maintained in an undifferentiated state via a tight transcriptional network. Additionally, other ESC-specific enhancers containing Oct3/4 and SOX2 binding sites have been identified in a number of genes, including FGF4 (Fibroblast Growth Factor-4), SPP1 (Secreted Phosphoprotein-1), UTF1 (Undifferentiated embryonic cell Transcription Factor-1), FBXO15 (F-box Protein-15), and Lefty1 (Left Right Determination Factor-1) (Ref.1 & 7). Numerous additional genes in Mouse have been identified recently that contain Oct4/SOX2 binding sites. Lig3 (Ligase-III, DNA, ATP-dependent), KCTD3 (Potassium Channel Tetramerisation Domain containing-3), BIN1 (Bridging Integrator-1), Bmi1 (Bmi1 Polycomb ring finger oncogene), and NASP (Nuclear Autoantigenic Sperm Protein (histone-binding)) are all genes that contain one Oct4/SOX2 binding site. InsIG2 (Insulin Induced Gene-2), Ipo11 (Importin-11), MYST4 (MYST Histone Acetyltransferase (monocytic leukemia)-4), NR6A1 (Nuclear Receptor Subfamily-6, Group A, Member-1), and STRBP are all examples of genes that contain 5 to 10 sites (Spermatid Perinuclear RNA Binding Protein). POU2F1 (POU domain, class 2, Transcription Factor-1), RANBP17 (RAN Binding Protein-17), and SMYD3 are all genes with more than ten sites (SET and MYND Domain containing-3). Oct4 is associated with 35 genes involved in chromatin remodeling. SMARCC1 (SWI/SNF related, Matrix associated, Actin dependent regulator of chromatin, Subfamily c, member-1), ARID1A (AT Rich Interactive Domain-1A (SWI-like)), ARID5B (AT Rich Interactive Domain-5B (SWI-like)), JARID1B (Jumonji, AT Rich Interactive Domain-1B), and JARID2 (Jumonji, AT Rich Interactive Domain-2) are putative positive target genes. These ARID domain-containing proteins, which comprise a subset of the Jumonji C family, have been recently linked to Histone Demethylase activity. Numerous additional genes have been identified that contain MYST, SET (SET translocation (Myeloid leukemia-associated)), Chromo, and Bromo domains that facilitate or recognize specific histone modifications. The ability of REST (RE1-Silencing Transcription Factor) to recruit cofactors such as HDACs (Histone Deacetylases), CoREST (Co-repressor of Rest), Sin3, and MECP2 has been implicated in the repression of neuronal specific genes (Methyl CpG binding Protein-2). The identification of REST as an Oct4 direct target, in light of its role in maintaining chromatin plasticity during neurogenesis, provides a mechanistic explanation for Oct4's role in neural differentiation. Oct4 is positively and negatively correlated with CoREST and MECP2, respectively
Learn about some of the most frequently asked question about Nanog, pluripotency, and embryonic stem cells.