This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
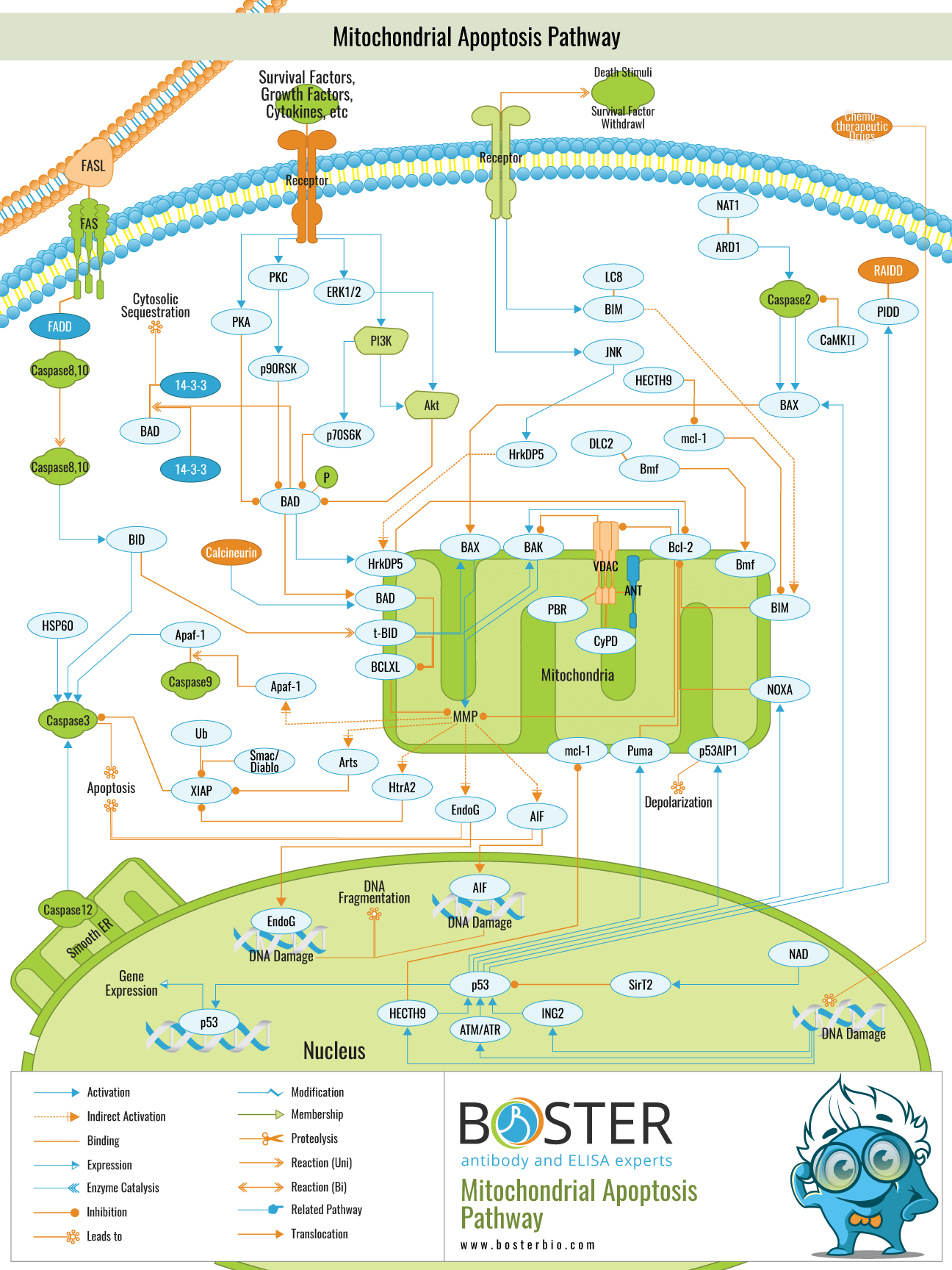
The endogenous mitochondrial pathway in cells can be activated by a death ligand or by apoptotic stimulating factors, resulting in cell death. Stimulating factors include oncogene activation, DNA damage, hypoxia, and deletion of cell growth factors, among others. In this pathway, the Bcl-2 family may be able to regulate the permeabilization of the mitochondrial outer membrane by adjusting the membrane potential. Special emphasis will be placed on novel aspects of the regulation of the Bcl-2 protein family's control of mitochondrial membrane permeabilization, including the mechanisms underlying their mitochondrial targeting and activation during apoptosis.
The function(s) of the oncosuppressive protein p53 at the mitochondria, and the role of mitochondrial fusion and fission processes.
The BCL-2 family proteins are apoptotic regulators, consisting of both anti- and pro-apoptotic members. They may regulate apoptosis by regulating mitochondrial outer membrane permeabilization (MOMP). The pro-apoptotic protein is classified into two subtypes: BH1-3 and BH3. Pro-apoptotic proteins such as Bak, as well as anti-apoptotic proteins such as Bcl-2 and Bcl-xL, are predominantly found in the mitochondrial membrane. Other members of the family, such as Bid and Bad, are predominantly found in the cytoplasm. Bax is normally found in the cytoplasm, but when activated by an apoptotic signal, it can migrate to the surface of mitochondria and form a transmembrane pore, lowering the membrane potential and increasing membrane permeability, which results in the release of apoptotic factor.
At the moment, two hypotheses exist regarding the activated modes of Bax and Bak: direct and indirect modes.Indirect mode: typically, anti-apoptotic proteins inhibit the activity of Bax and Bak, which can be activated in-directly if the anti-apoptotic activity is inhibited by members of the BH-3only family in response to an apoptotic signal. BH3-only can be classified as activated protein or kinase protein.
The activated proteins that have not been activated by an apoptotic signal may combine with anti-apoptotic proteins to inhibit Bax and Bak activation. By contrast, both Bax and Bak can be activated directly if the kinase protein combines with an anti-apoptotic protein, resulting in the release of activated protein upon receiving an apoptotic signal.
In conclusion, the Bcl-2 family contains anti-apoptotic and pro-apoptotic members that interact and jointly determine whether cells undergo apoptosis or not.
The decrease in the potential of the mitochondrial membrane and the increase in membrane permeability may result in the release of endogenous mitochondrial apoptotic factors such as CytC, AIF, SMAC/DIABLO, HTRA2/OMI, and ENDOG. After being released into cells, Cyt C interacts with Apaf-1 to form an apoptosis complex with the assistance of ATP and dATP. Then, the apoptosis complex combines with and activates Pro-Caspase9 to form Caspase9. Following that, Caspase9 could activate both Caspase3 and Caspase7, initiating the caspase cascade reaction and cleaving more than 100 different substrates in cells, including -tubulin, Actin, PARPA, and Lamin. Finally, the chain of events results in apoptosis.
IAPs may inhibit Caspase3 and Caspase7 activation, thereby inhibiting cell apoptosis. After egressing from the mitochondrion into the cytoplasm, SMAC/DIABLO and HTRA2/OMI may combine with IAPs to alleviate IAP-induced apoptosis inhibition, thereby indirectly promoting apoptosis.
AIF and ENDOG may also be released into the cytoplasm and then transferred into the cell nucleus, where they induce chromosome condensation and DNA fragmentation, ultimately resulting in apoptosis.