This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
Apoptosis or programmed cell death can be activated through the intrinsic death receptor pathway and the extrinsic death receptor pathway.
View pathway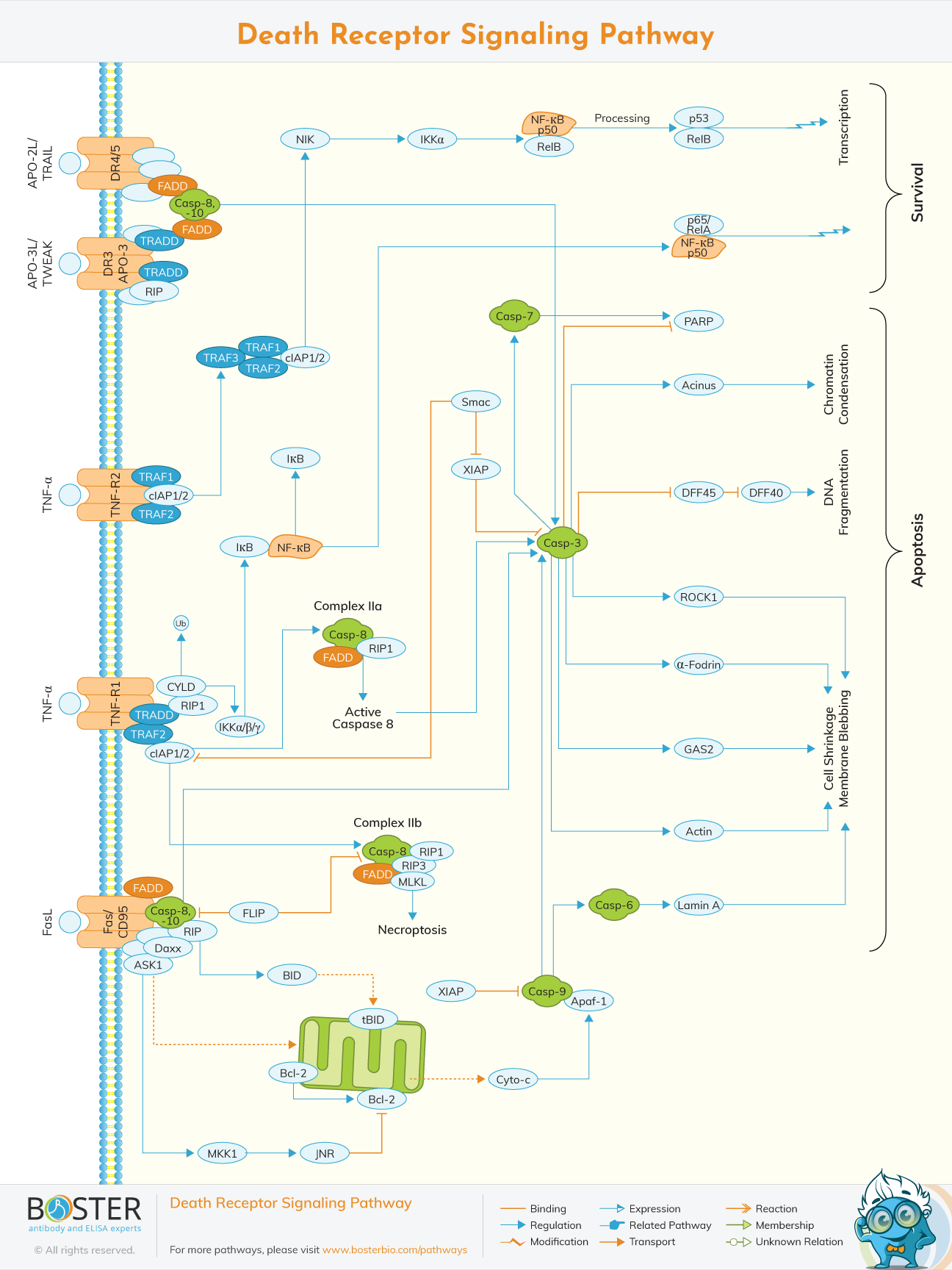
Apoptosis or programmed cell death can be activated through two distinct but related pathways. The first pathway, called the intrinsic death receptor pathway (aka intrinsic apoptosis pathway or mitochondrial pathway), is activated upon significant cell stress through the mitochondria while the second pathway, called the extrinsic death receptor pathway, is signaled by extracellular death ligands that bind to death receptors, triggering cell suicide. These death receptors are embedded in the cellular membrane and activate the apoptotic pathway when bound by their cognate death ligands. Following this, the death domains of the death receptors bind the TNFR-associated death domain (TRADD) which in turn recruits and interacts with the Fas-associated death domain (FADD). FADD then couples with procaspase 8 or 10, which are subsequently activated and cleaved to caspase 8 or 10, to form death-inducing signaling complexes (DISCs). These DISCs trigger the signal cascades implicated in apoptosis. Among the most well-known death receptors are Fas, TNFR1, TRAILR1, TRAILR2, DR3, and NGFR, which are all members of the tumor necrosis factor (TNF) receptor superfamily of proteins.
Apoptosis is a cellular process that has garnered significant interest in different biological research areas due to its role in development, immune response, and pathology of some human diseases, such as neurodegenerative diseases, autoimmune disorders, and most notably cancer. These disorders are all commonly characterized by defective apoptotic mechanisms in cells, which can be either impaired, as in the case of cancer, or excessive, as in the case of neurodegenerative diseases (Elmore, 2007). Thus, the proper signaling and modulation of the apoptosis pathway is necessary for normal physiological function, especially since apoptosis is a constantly occurring event in multicellular organisms that can be triggered by different stresses such as chemotherapy and irradiation (Lavrik, Golks, & Crammer, 2005). It is implicated in the elimination of undesired cells in the early development of organisms, while in adults it is involved in the removal of damaged cells (National Human Genome Research Institute, n.d.).
The term apoptosis was coined by Kerr, Wyllie, and Currie (1972) to refer to a form of cell death that differs in many ways to necrotic cell death. Changes in the morphological and biochemical makeup of cells precede apoptosis, including membrane blebbing, cell shrinkage, chromatin condensation, nuclear disruption, and DNA fragmentation. The cell cycle is also heavily involved in apoptosis, as cell cycle arrest events are required for apoptosis to initiate (Elmore 2007). While the linkage between cell cycle arrest and the death receptor pathway is faint, the pathway is heavily implicated in all the other morphological and biochemical cellular changes mentioned.
The elucidation of the death receptor pathway responsible for apoptosis have generated possible therapeutic targets in both the death receptors and their ligands (Kumar, Herbert, & Warrens, 2005). Each death receptor has a unique ligand that activates it and triggers the downstream signaling upon binding.
TNFR1/DR1 has TNF as its activating ligand, while Fas/DR2 has FasL as its cognate ligand. DR3/Apo3 has Apo3L/TWEAK as its activating ligand. The death ligand TRAIL/Apo2L is the cognate ligand of both TRAIL-R1/DR4 and TRAIL-R2/DR5. Lastly, DR6 has TRADD as its activating ligand that binds to its death domain instead of its extracellular domain. This is the case for all the other death receptors (Kumar, Herbert, & Warrens, 2005).
The death receptors are composed of extracellular cysteine-rich domains (CRDs) that bind the death ligands and a cytoplasmic death domain which recruits adapter molecules such as TRADD and FADD (Gurney, Muralidhar, & Linton, 2013). While all death receptors have a single intracellular death domain, the number of their extracellular CRDs differ.
TNFR1 or death receptor 1 (DR1), Apo3/DR3, and DR6 all have four CRDs in their extracellular domain. Fas/DR2 has three CRDs in its extracellular domain, while TRAIL-R1/DR4 and TRAIL-R2/DR5 both have two CRDs (Guicciardi & Gores, 2003). The death ligands bind to their cognate death receptors at the most distal CRD to the cellular membrane. Ultimately, the death receptor pathway is initiated by the activation of death receptors upon the binding of their cognate ligands.
In terms of genomic location, the genes coding for the death receptors are located on different chromosomes. TNFR1/DR1 can be found in chromosome 12p13, while Fas/DR2 is located in chromosome 10q23. Apo3/DR3 is located in chromosome 1p36 while both TRAIL-R1/DR4 and TRAIL-R2/DR5 are found in chromosome 8p21. Lastly, DR6 can be found in chromosome 6p12. Like their cognate receptors, the genomic locations of the death ligands also vary widely. TNF is situated in chromosome 6p21 while FasL can be found in chromosome 1q24. Apo3L/TWEAK is located in chromosome 17p13 while TRAIL/Apo2L can be found in chromosome 3q26. Lastly, TRADD is located in chromosome 16q22. Interestingly, none of the death receptors are located in the same chromosome, as is true for the death ligands as well.
Proteins in the death receptor pathway, especially the death receptors themselves, serve as valuable therapeutic targets as a result of both their cytotoxic and non-cytotoxic effects. The specificity of death ligands to their cognate death receptors have made them valuable molecular targets for the targeted therapy of diseases involving the death receptor pathway (Sheikh & Huang, 2004).
Fas, the most studied of the death receptors, has also emerged as one of the top drug targets for therapies involving the death receptor pathway. Known anticancer drugs such as doxorubicin, cisplatin, cytarabine, and methotrexate have been shown by Friesen, Fulsa, & Debatin (1999) to induce FasL and increase its binding to Fas, resulting in increased cell death events. Thus, one of the mechanisms by which these drugs promote tumor elimination is by promoting apoptosis mediated by Fas signaling. Tumor cells resistant to chemotherapies have also been studied to have defective Fas signaling. Antibodies that bind the death receptors have been shown to reduce apoptosis by blocking the binding of death ligands as well. This is in turn helpful in pathologies where there may be excessive apoptosis, such as neurodegenerative diseases.
The death receptor pathway influences many diseases and pathologies, rendering death receptors, death ligands, and other proteins in the pathway as valuable therapeutic targets that can be investigated through immunoassays such as ELISA. Boster helps with research on death receptor signaling by offering a wide range of antibodies that reliably target proteins in the death receptor pathway.