This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
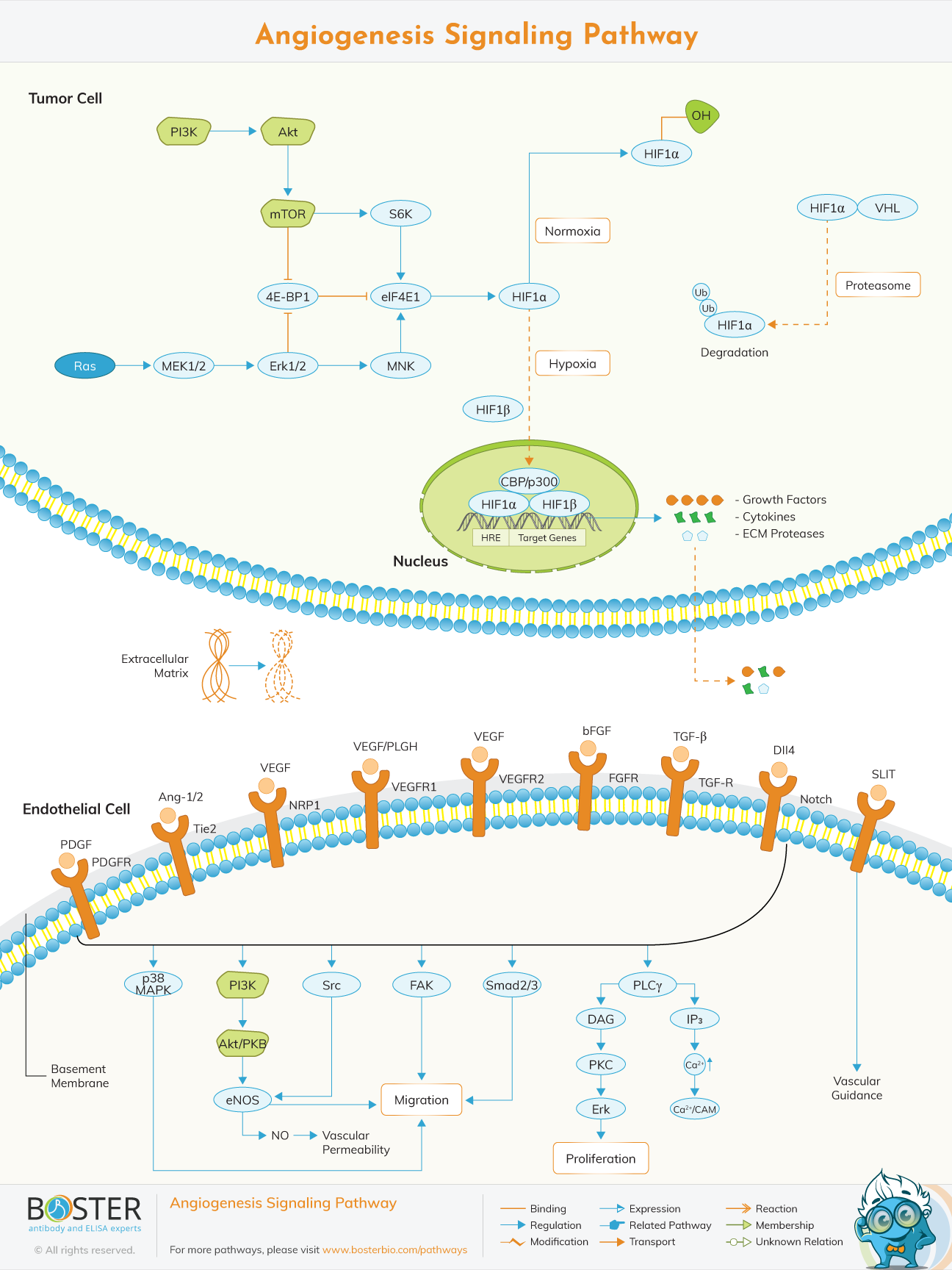
Angiogenesis is the process by which new blood vessels are formed from vasculature due to different stimuli. Angiogenesis is a complicated and well-ordered process that involves extensive signaling networks between and inside endothelial cells (ECs), mural cells (VSMCs and pericytes), and other cell types (eg, immune cells).
Angiogenesis is mediated primarily by VEGF. VEGF (vascular endothelial growth factor) is a protein family that is necessary for angiogenesis. VEGF comes in a variety of types, including VEGF-A, VEGF-B, VEGF-C, and VEGF-D, each of which plays a key role in angiogenic processes ranging from embryonic to lymphatic angiogenesis.
There are two main types of angiogenesis Sprouting Angiogenesis and Intussusceptive Angiogenesis.
Sprouting angiogenesis is characterized by endothelial cell sprouts that typically develop toward an angiogenic stimulus like VEGF-A.
Intussusceptive angiogenesis involves formation of blood vessels by a splitting process in which elements of interstitial tissues invade existing vessels, forming transvascular tissue pillars that expand.
Hypoxia, which is normal in tumors, is one of the most powerful triggers for increased VEGF production by tumor cells. Hypoxia causes hypoxia-inducible factor 1 (HIF-1), a bHLH–PAS transcription factor with both a and a subunit, to be activated.HIF-1α is upregulated across a broad range of cancers, and HIF regulates key features of tumor biology such as angiogenesis, invasion, and glucose metabolism.
Though hypoxia is the most common cause of HIF-1 upregulation, activation of the epidermal growth factor receptor (EGFR) and the PI3K pathway can also cause HIF-1 to increase. EGFR is a tyrosine kinase transmembrane receptor that belongs to the HER family of receptors. RAS mutation, increased expression of growth factor receptors such as EGFR, or loss of PTEN may all cause PI3K activation. The PI3K/AKT/mTOR pathway can be activated to increase VEGF secretion. Other angiogenic factors including nitric oxide and angiopoietins are also influenced by the PI3K/AKT pathway.
H-RAS, N-RAS, and K-RAS are members of the RAS family of GTPases, which are involved in a variety of cellular functions such as proliferation, differentiation, and apoptosis.RAS upregulates Glut1 promoter activity, at least in part, by increasing HIF-1α protein and that this involves the PI3K pathway.
The PI3K pathway also plays an essential role in the formation of normal blood vessels during development. Embryos with kinase-dead p110α catalytic subunit of PI3K develop gross vascular defects, indicating that this subunit is essential for endothelial cell migration and angiogenesis. When VEGF binds to its receptor on normal endothelial cells, the RAS and PI3K pathways are activated. Constitutive activation of the ERK/MAPK pathway could induce angiogenesis but not vascular permeability, while activation of PI3K was required for both angiogenesis and vascular permeability. Slit and Robo proteins are vascular remodeling regulators. The mTORC2-dependent activation of AKT and RAC by Slit 2 protein will promote angiogenesis. However, in the presence of Ephrin A1, this mechanism is inhibited, implying that Ephrin A1-Slit signaling regulates angiogenesis and vascular homeostasis.
Via its control of nitric oxide (NO) signaling in endothelial cells, the PI3K/AKT pathway has also been linked to angiogenesis. The enzyme NO synthase controls nitric oxide synthesis (NOS). NOS has three isoforms: nNOS/NOS1, eNOS/NOS3, and iNOS/NOS2. nNOS/NOS1 and eNOS/NOS3 are constitutively expressed, while iNOS/NOS2 expression is inducible.
Both HIF-1 dependent and independent mechanisms will increase VEGF secretion when the PI3K/AKT/mTOR pathway is activated in tumor cells. By modulating the expression of nitric oxide and angiopoietins, this pathway can also control angiogenesis.
John Hunter, a Scottish anatomist and surgeon, was the first to report scientific findings in the field of angiogenesis. His findings indicated that vascularity and metabolic requirements are proportional in both health and disease. This conviction is summarized in his Treatise, which was published in 1794.
Judah Folkman, who hypothesized (and published in 1971) that tumor growth is based on angiogenesis, is credited with starting the modern history of angiogenesis. The realization that controlling angiogenesis could lead to cancer therapies sparked a flurry of activity in the field; for example, in 1970, only two manuscripts dealing with angiogenesis were published; by 2009, over 5200 articles had been published.
The discovery of the molecular mechanisms of physiological vasculogenesis and pathological angiogenesis aided in the identification of two classes of diseases: those in which therapeutic angiogenesis can repair tissue damage (arteriosclerosis, myocardial infarction, limb ischemia) and those in which pathological angiogenesis inhibition can cure or delay disease progression (retinopathies, benign and malignant angiogenic tumors, progression of malignant tumors)
TIn healthy tissues, the body regulates angiogenesis by establishing a perfect balance of growth and inhibitory substances. When this equilibrium is upset, either too much or too little angiogenesis occurs. Abnormal blood vessel development, whether excessive or inadequate, is increasingly recognized as a “common denominator” underlying a wide range of fatal and disabling illnesses.
The use of biological agents, bioactive materials, or environmental factors to encourage the creation of new vessels, restore or enhance circulatory perfusion of tissues, reverse ischemia, or expedite healing is known as therapeutic angiogenesis. This method is presently employed in the treatment of heart-cerebrovascular disorders, critical limb ischemia, delayed wound healing, and peptic ulcer disease.
Biocompatible materials including matrices and angiogenesis modulators can be utilized to improve healing in cases of serious wounds and skin losses such as burns. Healing can be aided with a wound dressing containing a synthetic copper-peptide that may collect FGF. Another approach is to utilize a tissue-engineered biological covering containing cells that generate huge amounts of angiogenic factors such as FGF or VEGF.
A laser beam can induce transmyocardial revascularization in cases of cardiac ischemia. Myocytes, inflammatory cells, and platelets offer a substantial supply of angiogenic factors following photoacustic damage.
This gadget is currently designed for both intraoperative (bypass surgery) and transcutaneous usage.
The primary purpose of hyperbaric oxygenation is to heal superficial wounds. Patients are placed in a chamber where the oxygen pressure is gradually elevated to much above the physiological level. A systemic oxygen burst occurs when blood plasma begins to transport an increasing amount of oxygen. It has been demonstrated that elevated oxygen tension enhances endothelial cell growth and causes angiogenesis in ischemic tissues.
Several angiogenic growth factors are now being studied in clinical trials, including the most potent and ubiquitous ones like VEGF, a/bFGF, PDGF, and G-CSF. These experiments use human recombinant proteins, gene technology for delivering bare DNA, and adenoviral vectors to transfect endothelial cells. The clinical use of G-CSF is based on the observation that contrary to what was thought, the embryonic type of angiogenesis (mobilization of bone marrowendothelial stem cells to the site of neoangiogenesis) is possible in adults as well, since endothelial stem cells are continously present, though in a low number, in the circulation.