This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

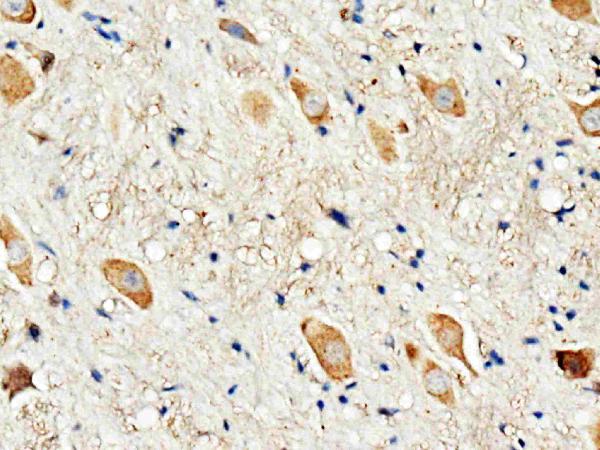
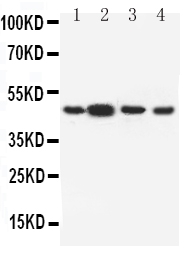

Facts about D(3) dopamine receptor.

.
| Human | |
|---|---|
| Gene Name: | DRD3 |
| Uniprot: | P35462 |
| Entrez: | 1814 |

| Belongs to: |
|---|
| G-protein coupled receptor 1 family |

D(3) dopamine receptor; D3DR; Dopamine D3 R; Dopamine D3 receptor; Dopamine D3R; dopamine receptor D3; DRD3; essential tremor 1; ETM1; FET1; MGC149204; MGC149205
Mass (kDA):
44.225 kDA

| Human | |
|---|---|
| Location: | 3q13.31 |
| Sequence: | 3; NC_000003.12 (114127580..114199407, complement) |
Brain.
Cell membrane; Multi-pass membrane protein. Both membrane-bound and scattered in the cytoplasm during basal conditions. Receptor stimulation results in the rapid internalization and sequestration of the receptors at the perinuclear area (5 and 15 minutes), followed by the dispersal of the receptors to the membrane (30 minutes). DRD3 and GRK4 co-localize in lipid rafts of renal proximal tubule cells.



PMID: 2129115 by Giros B., et al. Gene cloning of human dopaminergic D3 receptor and identification of its chromosome.
PMID: 8415635 by Schmauss C., et al. Selective loss of dopamine D3-type receptor mRNA expression in parietal and motor cortices of patients with chronic schizophrenia.