This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

2 Citations 5 Q&As


2 Citations 17 Q&As
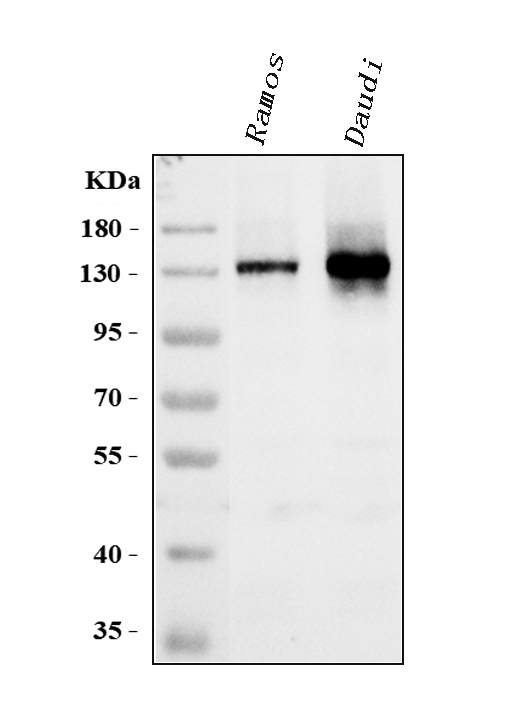











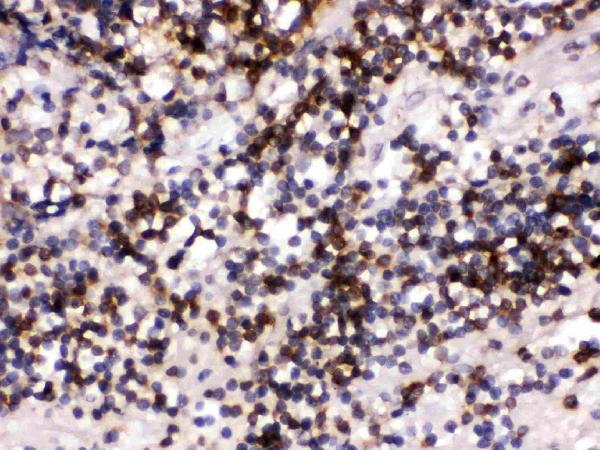
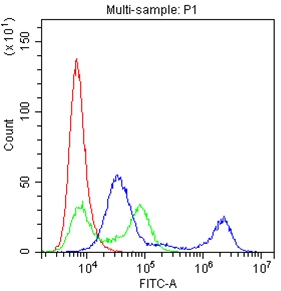
Facts about B-cell receptor CD22.

Preferentially binds to alpha-2,6-linked sialic acid. The sialic acid recognition site could be masked by cis interactions with sialic acids on the same cell surface.
| Human | |
|---|---|
| Gene Name: | CD22 |
| Uniprot: | P20273 |
| Entrez: | 933 |

| Belongs to: |
|---|
| immunoglobulin superfamily |

B-cell receptor CD22; BL-CAM; B-lymphocyte cell adhesion molecule; CD22 antigenMGC130020; CD22 molecule; CD22; sialic acid binding Ig-like lectin 2; Sialic acid-binding Ig-like lectin 2; Siglec2; Siglec-2; SIGLEC2FLJ22814; T-cell surface antigen Leu-14
Mass (kDA):
95.348 kDA

| Human | |
|---|---|
| Location: | 19q13.12 |
| Sequence: | 19; NC_000019.10 (35329169..35347361) |
B-lymphocytes.
Cell membrane; Single-pass type I membrane protein.





PMID: 1691828 by Stamenkovic I., et al. The B-cell antigen CD22 mediates monocyte and erythrocyte adhesion.
PMID: 1985119 by Wilson G.L., et al. cDNA cloning of the B cell membrane protein CD22: a mediator of B-B cell interactions.
*More publications can be found for each product on its corresponding product page









