This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

30 Citations 5 Q&As
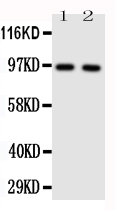
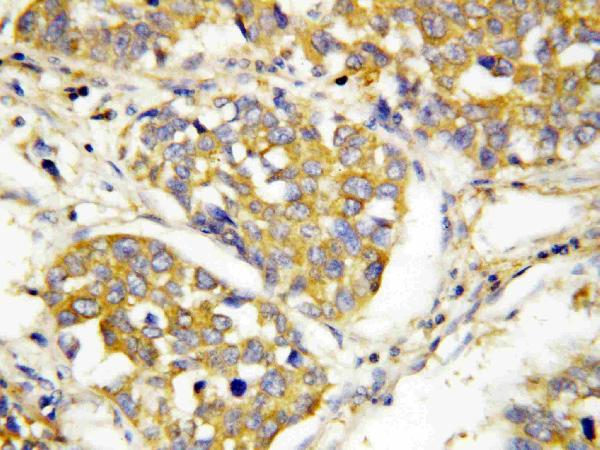
12 Citations 16 Q&As


Facts about Toll-like receptor 4.

Responses triggered by Ni(2+) require non-conserved histidines and are, therefore, species-specific (PubMed:20711192). Both M.
| Human | |
|---|---|
| Gene Name: | TLR4 |
| Uniprot: | O00206 |
| Entrez: | 7099 |

| Belongs to: |
|---|
| Toll-like receptor family |

76B357.1; ARMD10; CD284 antigen; CD284; hTollhomolog of Drosophila toll; tlr4 76B357.1; TLR4 facs; TLR4 flow cytometry; TLR4 human; TLR4 ihc; TLR4 western blot; TLR4; TOLL; toll-like receptor 4
Mass (kDA):
95.68 kDA

| Human | |
|---|---|
| Location: | 9q33.1 |
| Sequence: | 9; NC_000009.12 (117704403..117724735) |
Highly expressed in placenta, spleen and peripheral blood leukocytes (PubMed:9435236, PubMed:9237759). Detected in monocytes, macrophages, dendritic cells and several types of T-cells (PubMed:9237759, PubMed:27022195).
Cell membrane; Single-pass type I membrane protein. Early endosome. Cell projection, ruffle. Upon complex formation with CD36 and TLR6, internalized through dynamin-dependent endocytosis (PubMed:20037584). Colocalizes with RFTN1 at cell membrane and then together with RFTN1 moves to endosomes, upon lipopolysaccharide stimulation.





PMID: 9237759 by Medzhitov R., et al. A human homologue of the Drosophila Toll protein signals activation of adaptive immunity.
PMID: 9435236 by Rock F.L., et al. A family of human receptors structurally related to Drosophila Toll.
Tetramethylpyrazine attenuates endotoxin-induced retinal inflammatio...
*Showing only the more recent 20. More publications can be found for each product on its corresponding product page




